

Molecules incorporating a benzothiazole core scaffold inhibit the N-myristoyltransferase of Plasmodium falciparum
- PMID: 17714074
- PMCID: PMC2267354
- DOI: 10.1042/BJ20070692
Molecules incorporating a benzothiazole core scaffold inhibit the N-myristoyltransferase of Plasmodium falciparum
Abstract
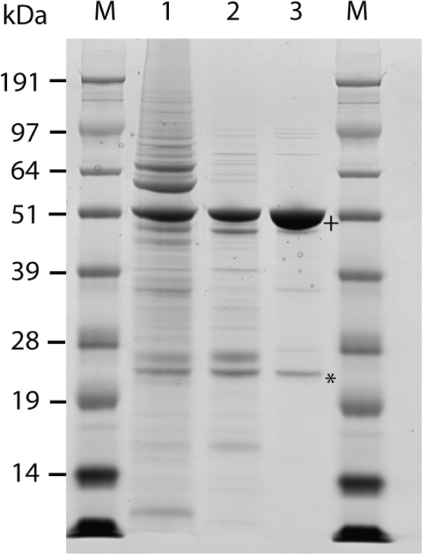
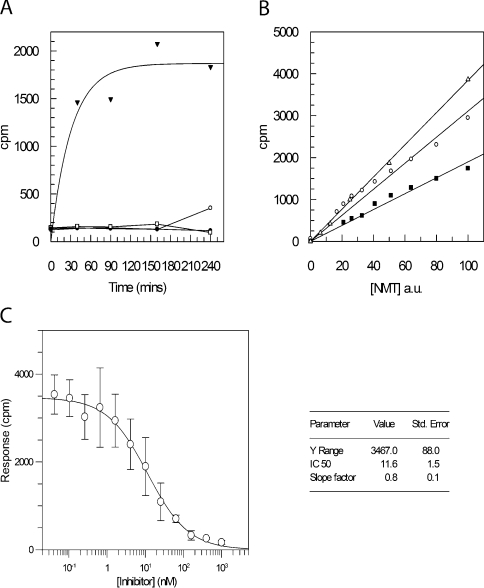
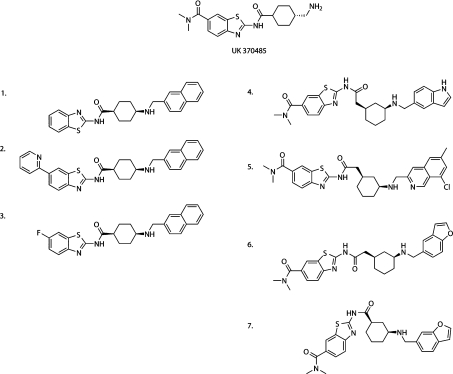
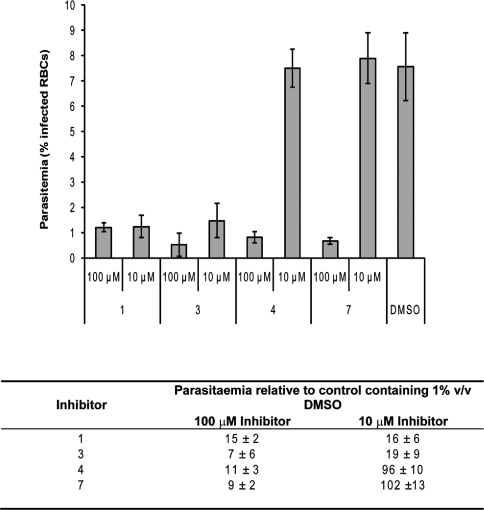
Recombinant N-myristoyltransferase of Plasmodium falciparum (termed PfNMT) has been used in the development of a SPA (scintillation proximity assay) suitable for automation and high-throughput screening of inhibitors against this enzyme. The ability to use the SPA has been facilitated by development of an expression and purification system which yields considerably improved quantities of soluble active recombinant PfNMT compared with previous studies. Specifically, yields of pure protein have been increased from 12 microg x l(-1) to >400 microg x l(-1) by use of a synthetic gene with codon usage optimized for expression in an Escherichia coli host. Preliminary small-scale 'piggyback' inhibitor studies using the SPA have identified a family of related molecules containing a core benzothiazole scaffold with IC50 values <50 microM, which demonstrate selectivity over human NMT1. Two of these compounds, when tested against cultured parasites in vitro, reduced parasitaemia by >80% at a concentration of 10 microM.
Figures
References
-
- Wilcox C., Hu J. S., Olson E. N. Acylation of proteins with myristic acid occurs cotranslationally. Science. 1987;238:1275–1278. - PubMed
-
- Resh M. D. Fatty acylation of proteins: new insights into membrane targeting of myristoylated and palmitoylated proteins. Biochim. Biophys. Acta. 1999;1451:1–16. - PubMed
-
- Maurer-Stroh S., Eisenhaber B., Eisenhaber F. N-terminal N-myristoylation of proteins: refinement of the sequence motif and its taxon-specific differences. J. Mol. Biol. 2002;317:523–540. - PubMed
-
- Maurer-Stroh S., Eisenhaber F. Myristoylation of viral and bacterial proteins. Trends Microbiol. 2004;12:178–185. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources