

Sensory adaptation
- PMID: 17714934
- PMCID: PMC2084204
- DOI: 10.1016/j.conb.2007.07.001
Sensory adaptation
Abstract
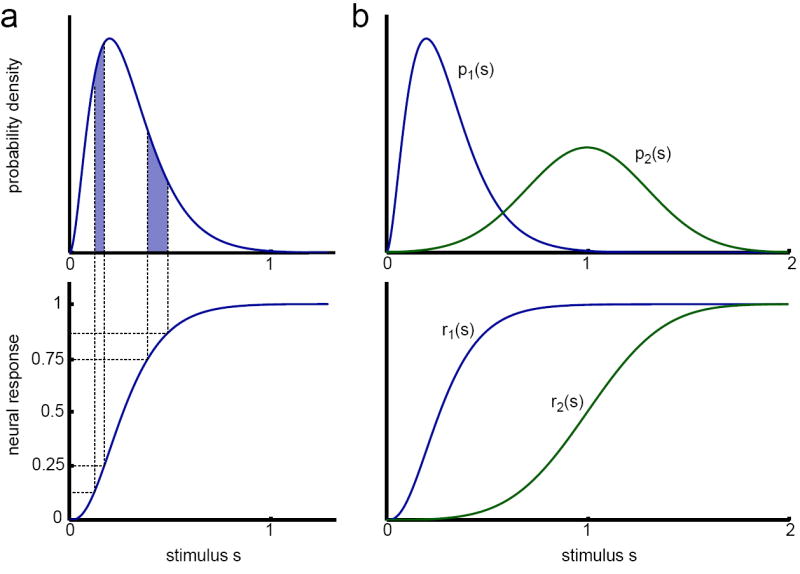
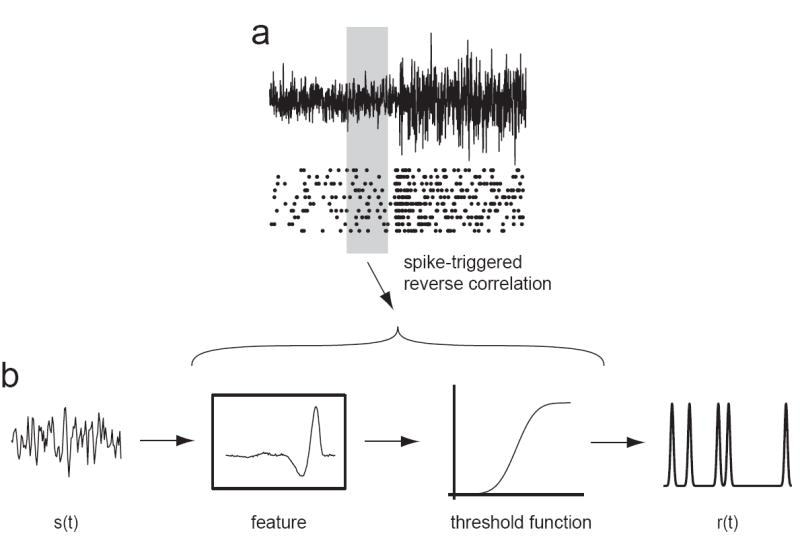
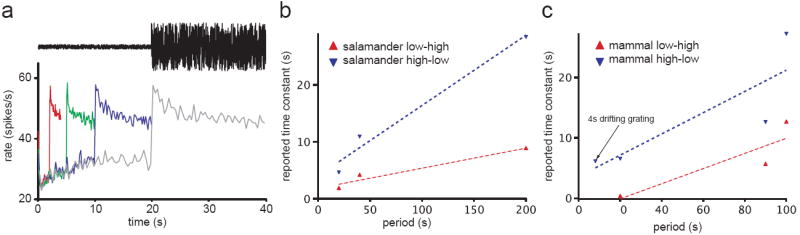
Adaptation occurs in a variety of forms in all sensory systems, motivating the question: what is its purpose? A productive approach has been to hypothesize that adaptation helps neural systems to efficiently encode stimuli whose statistics vary in time. To encode efficiently, a neural system must change its coding strategy, or computation, as the distribution of stimuli changes. Information theoretic methods allow this efficient coding hypothesis to be tested quantitatively. Empirically, adaptive processes occur over a wide range of timescales. On short timescales, underlying mechanisms include the contribution of intrinsic nonlinearities. Over longer timescales, adaptation is often power-law-like, implying the coexistence of multiple timescales in a single adaptive process. Models demonstrate that this can result from mechanisms within a single neuron.
Figures
References
-
- Simoncelli EP. Vision and the statistics of the visual environment. Curr Opin Neurobiol. 2003;13:144–149. - PubMed
-
- Barlow HB. Possible principles underlying the transformation of sensory messages. In: Rosenblith W, editor. Sensory Communication. MIT Press; 1961.
-
- Laughlin SB. The role of sensory adaptation in the retina. J Exp Biol. 1989;146:39–62. - PubMed
-
- Ulanovsky N, Las L, Nelken I. Processing of low-probability sounds by cortical neurons. Nat Neurosci. 2003;6:391–398. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
