

Requirement for Slit-1 and Robo-2 in zonal segregation of olfactory sensory neuron axons in the main olfactory bulb
- PMID: 17715346
- PMCID: PMC6672192
- DOI: 10.1523/JNEUROSCI.2217-07.2007
Requirement for Slit-1 and Robo-2 in zonal segregation of olfactory sensory neuron axons in the main olfactory bulb
Abstract
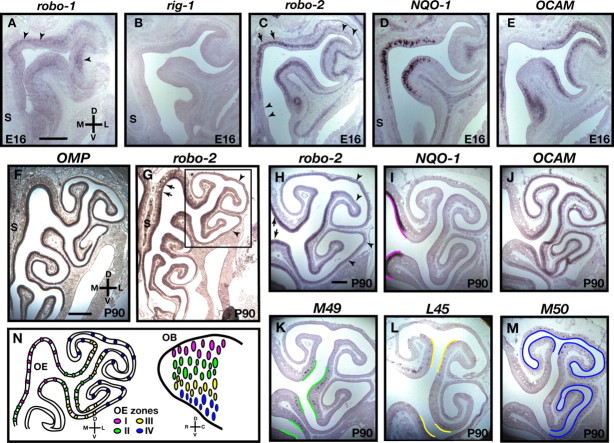
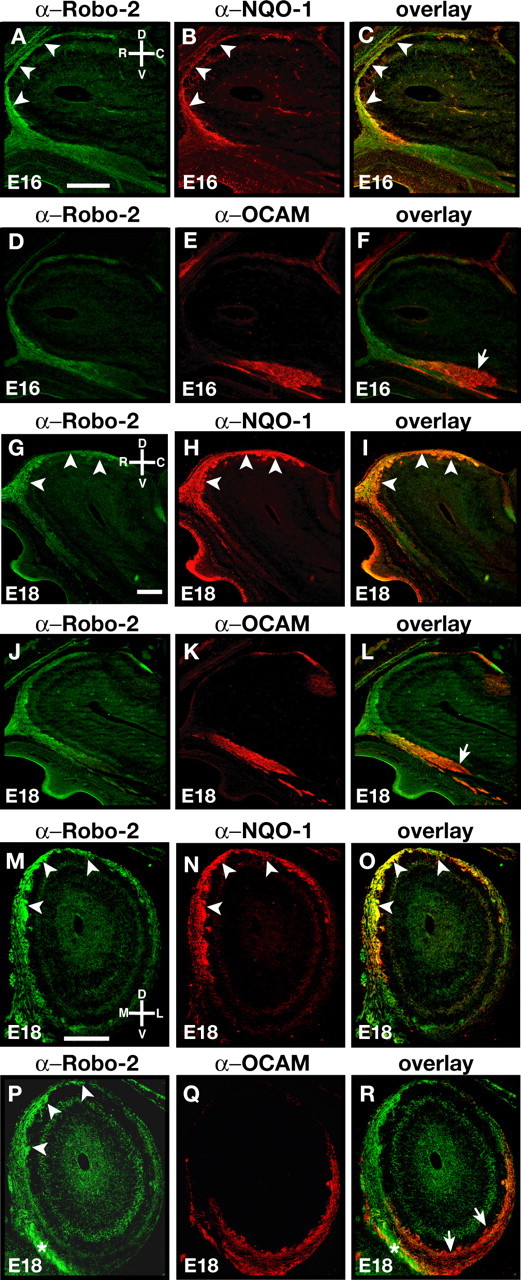
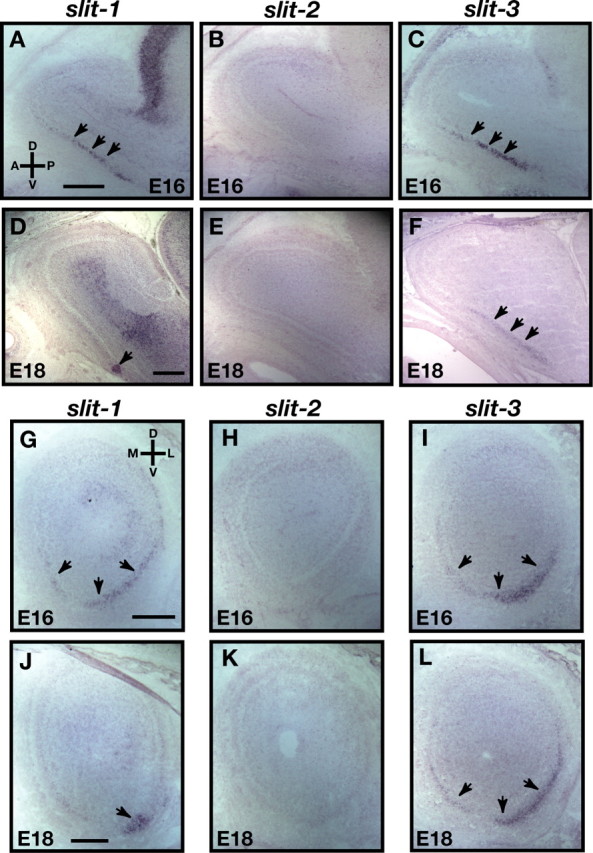
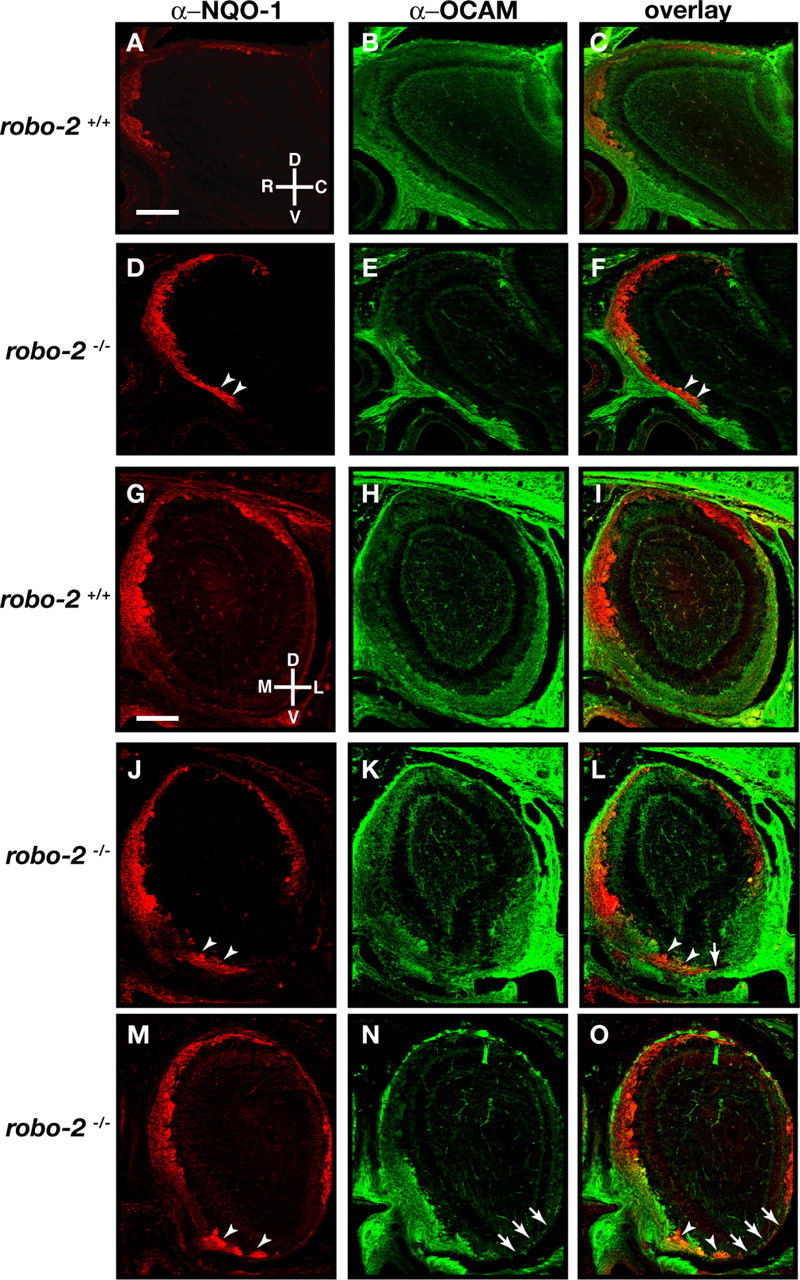
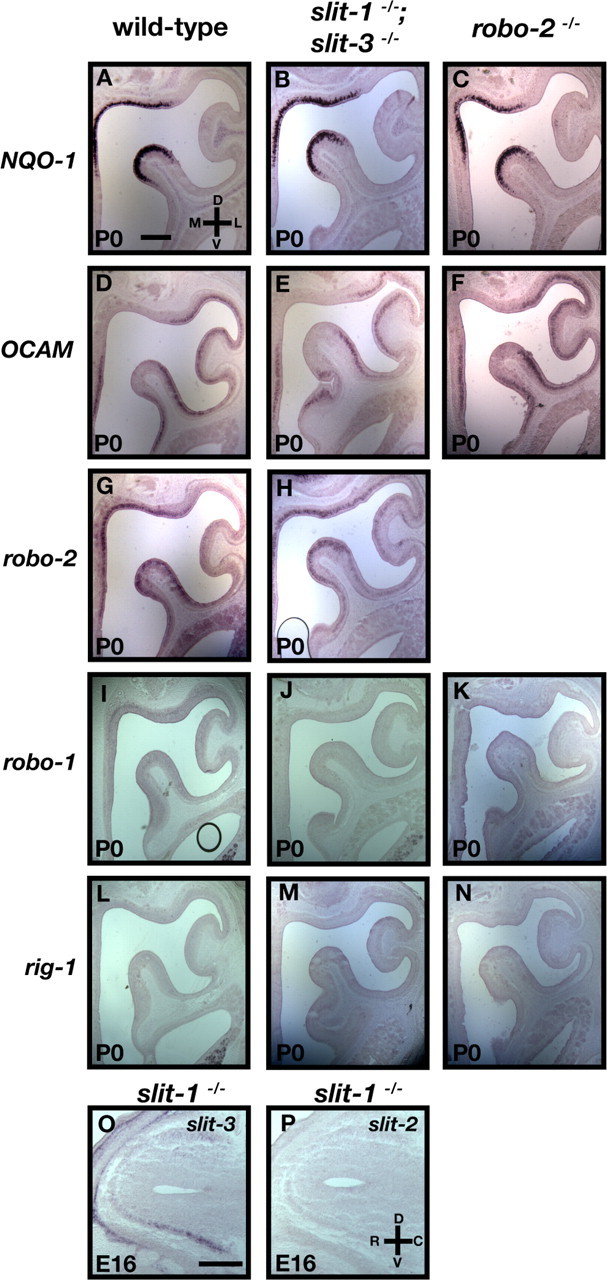
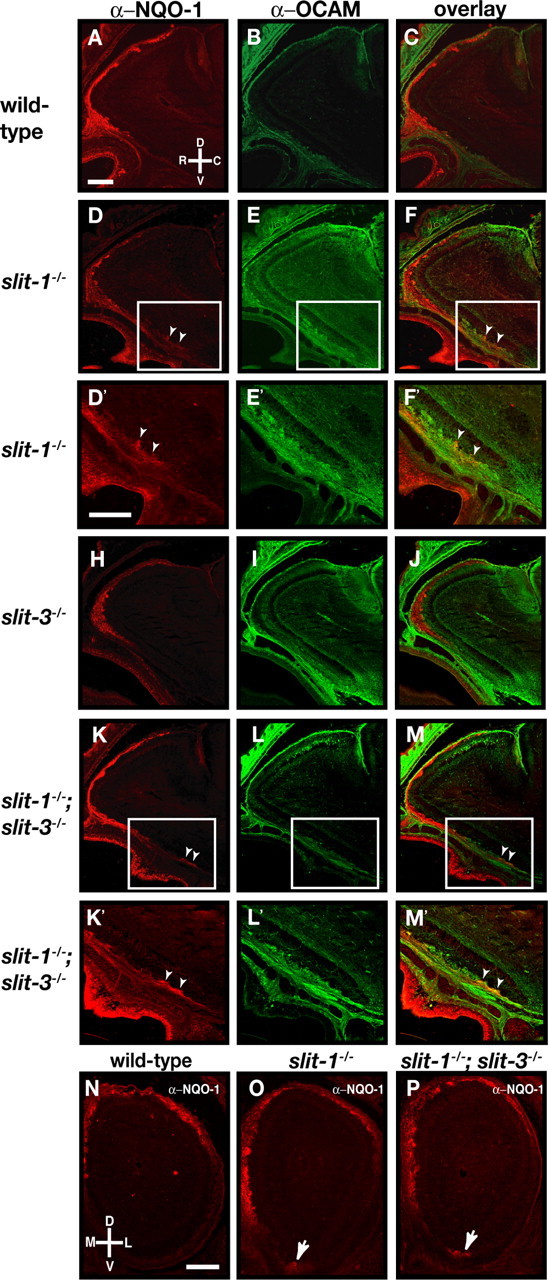
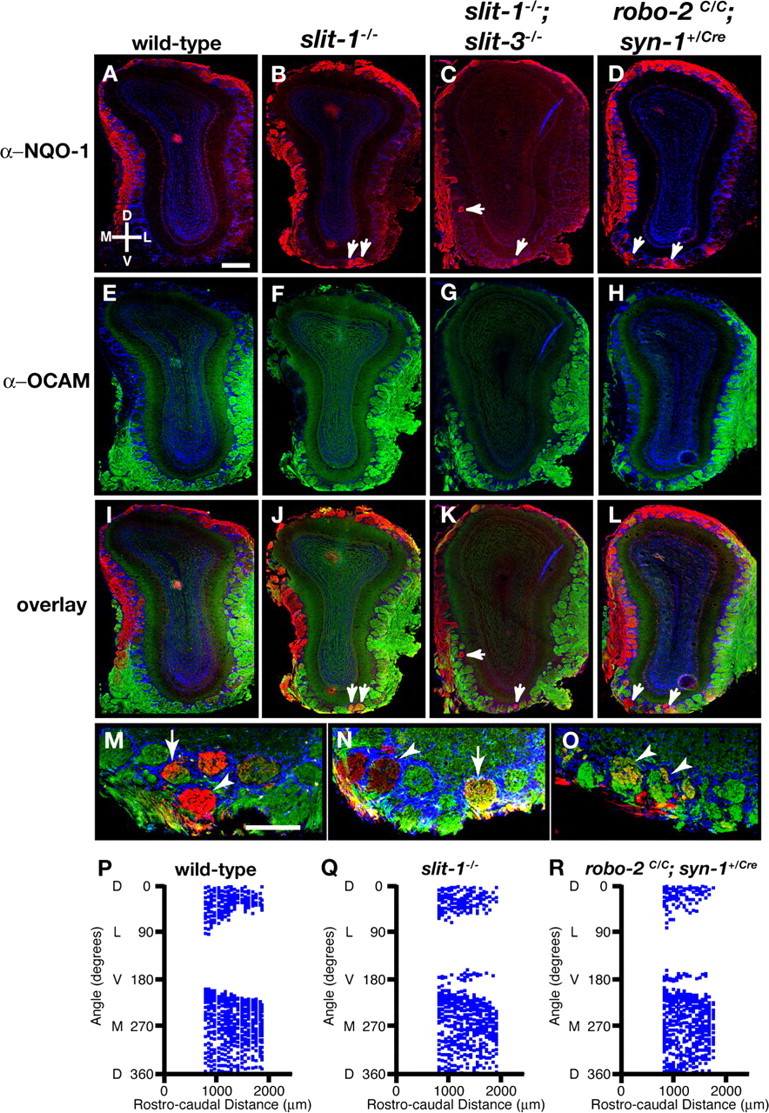
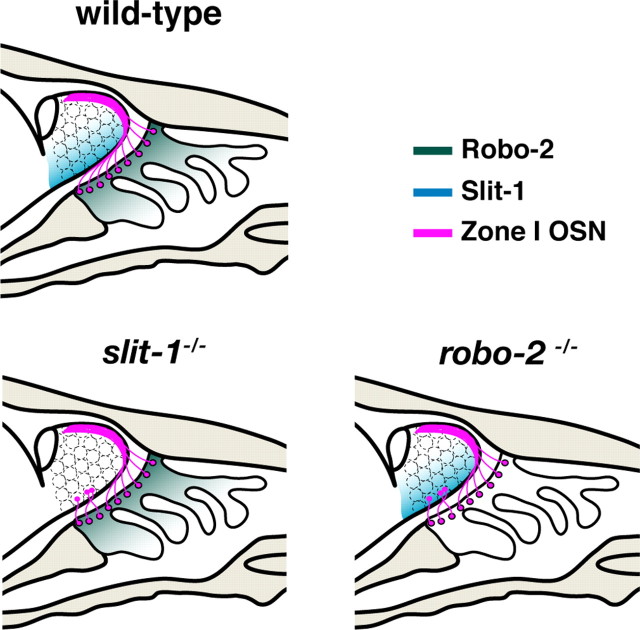
The formation of precise stereotypic connections in sensory systems is critical for the ability to detect and process signals from the environment. In the olfactory system, olfactory sensory neurons (OSNs) project axons to spatially defined glomeruli within the olfactory bulb (OB). A spatial relationship exists between the location of OSNs within the olfactory epithelium (OE) and their glomerular targets along the dorsoventral axis in the OB. The molecular mechanisms underlying the zonal segregation of OSN axons along the dorsoventral axis of the OB are poorly understood. Using robo-2(-/-) (roundabout) and slit-1(-/-) mice, we examined the role of the Slit family of axon guidance cues in the targeting of OSN axons during development. We show that a subset of OSN axons that normally project to the dorsal region of the OB mistarget and form glomeruli in the ventral region in robo-2(-/-) and slit-1(-/-) mice. In addition, we show that the Slit receptor, Robo-2, is expressed in OSNs in a high dorsomedial to low ventrolateral gradient across the OE and that Slit-1 and Slit-3 are expressed in the ventral region of the OB. These results indicate that the dorsal-to-ventral segregation of OSN axons are not solely defined by the location of OSNs within the OE but also relies on axon guidance cues.
Figures
Similar articles
-
Slits and Robo-2 regulate the coalescence of subsets of olfactory sensory neuron axons within the ventral region of the olfactory bulb.Dev Biol. 2012 Nov 15;371(2):269-79. doi: 10.1016/j.ydbio.2012.08.028. Epub 2012 Sep 7. Dev Biol. 2012. PMID: 22981605
-
Differential timing of neurogenesis underlies dorsal-ventral topographic projection of olfactory sensory neurons.Neural Dev. 2017 Feb 13;12(1):2. doi: 10.1186/s13064-017-0079-0. Neural Dev. 2017. PMID: 28193234 Free PMC article.
-
Inverse expression of olfactory cell adhesion molecule in a subset of olfactory axons and a subset of mitral/tufted cells in the developing rat main olfactory bulb.J Comp Neurol. 2003 Apr 14;458(4):389-403. doi: 10.1002/cne.10590. J Comp Neurol. 2003. PMID: 12619073
-
Axon guidance events in the wiring of the mammalian olfactory system.Mol Neurobiol. 2009 Feb;39(1):1-9. doi: 10.1007/s12035-008-8047-7. Epub 2008 Dec 2. Mol Neurobiol. 2009. PMID: 19048417 Review.
-
Developmental regulation of neural map formation in the mouse olfactory system.Dev Neurobiol. 2015 Jun;75(6):594-607. doi: 10.1002/dneu.22268. Epub 2015 Feb 18. Dev Neurobiol. 2015. PMID: 25649346 Review.
Cited by
-
IgSF8: a developmentally and functionally regulated cell adhesion molecule in olfactory sensory neuron axons and synapses.Mol Cell Neurosci. 2012 Jul;50(3-4):238-49. doi: 10.1016/j.mcn.2012.05.007. Epub 2012 Jun 9. Mol Cell Neurosci. 2012. PMID: 22687584 Free PMC article.
-
Spatial transcriptomic reconstruction of the mouse olfactory glomerular map suggests principles of odor processing.Nat Neurosci. 2022 Apr;25(4):484-492. doi: 10.1038/s41593-022-01030-8. Epub 2022 Mar 21. Nat Neurosci. 2022. PMID: 35314823 Free PMC article.
-
Wiring the senses: Factors that regulate peripheral axon pathfinding in sensory systems.Dev Dyn. 2023 Jan;252(1):81-103. doi: 10.1002/dvdy.523. Epub 2022 Aug 30. Dev Dyn. 2023. PMID: 35972036 Free PMC article. Review.
-
The pattern of glomerular map formation defines responsiveness to aversive odorants in mice.J Neurosci. 2011 May 25;31(21):7920-6. doi: 10.1523/JNEUROSCI.2460-10.2011. J Neurosci. 2011. PMID: 21613506 Free PMC article.
-
Coordination of olfactory receptor choice with guidance receptor expression and function in olfactory sensory neurons.PLoS Genet. 2018 Jan 31;14(1):e1007164. doi: 10.1371/journal.pgen.1007164. eCollection 2018 Jan. PLoS Genet. 2018. PMID: 29385124 Free PMC article.
References
-
- Alenius M, Bohm S. Identification of a novel neural cell adhesion molecule-related gene with a potential role in selective axonal projection. J Biol Chem. 1997;272:26083–26086. - PubMed
-
- Astic L, Saucier D. Anatomical mapping of the neuroepithelial projection to the olfactory bulb in the rat. Brain Res Bull. 1986;16:445–454. - PubMed
-
- Brose K, Bland KS, Wang KH, Arnott D, Henzel W, Goodman CS, Tessier-Lavigne M, Kidd T. Slit proteins bind Robo receptors and have an evolutionarily conserved role in repulsive axon guidance. Cell. 1999;96:795–806. - PubMed
-
- Cloutier J-F, Giger RJ, Koentges G, Dulac C, Kolodkin AL, Ginty DD. Neuropilin-2 mediates axonal fasciculation, zonal segregation, but not axonal convergence, of primary accessory olfactory neurons. Neuron. 2002;33:877–892. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous