

Alcohol ingestion by donors amplifies experimental airway disease after heterotopic transplantation
- PMID: 17717204
- PMCID: PMC2176096
- DOI: 10.1164/rccm.200702-255OC
Alcohol ingestion by donors amplifies experimental airway disease after heterotopic transplantation
Abstract
Rationale: Obliterative bronchiolitis (OB) after lung transplantation is triggered by alloimmunity, but is ultimately mediated by transforming growth factor (TGF)-beta(1)-dependent airway fibrosis.
Objectives: Chronic alcohol use increases TGF-beta(1) expression and renders the lung susceptible to injury. Therefore, we hypothesized that donor alcohol abuse could prime the lung allograft for OB, as many organ donors have a history of alcohol abuse.
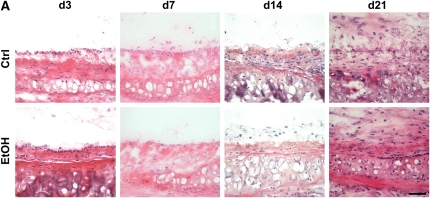
Methods: Tracheas from control and alcohol-fed rats (8 wk) were heterotopically transplanted into recipients with varying degrees of alloimmune mismatch and analyzed for obliterative airway disease severity on Postoperative Day 21.
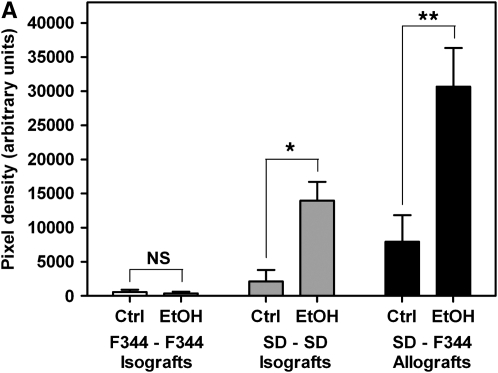
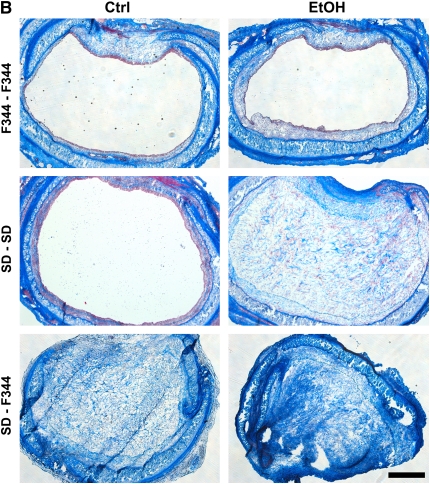
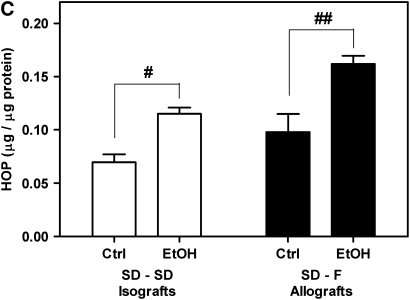
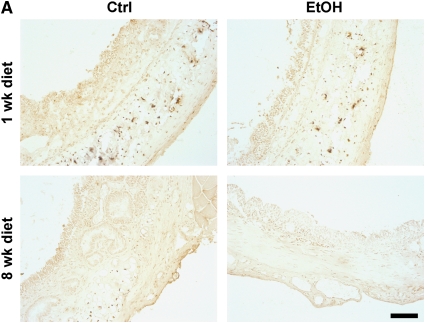
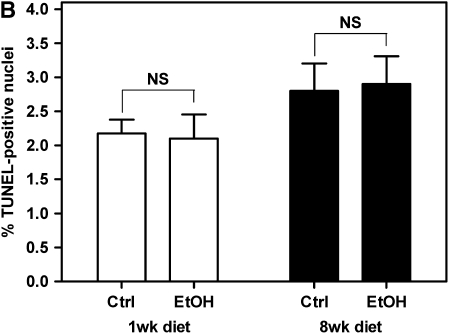
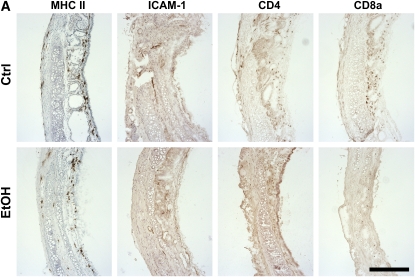
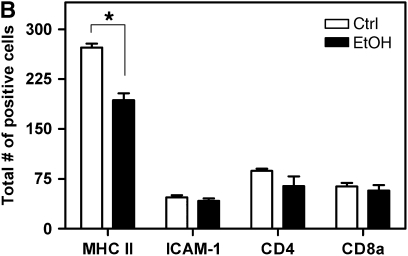
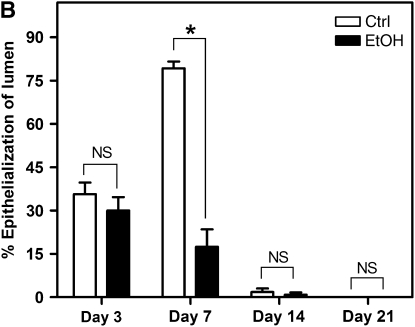
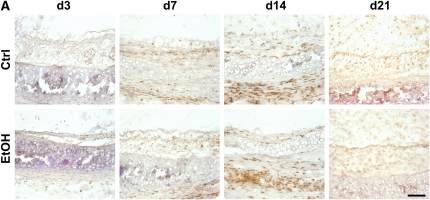
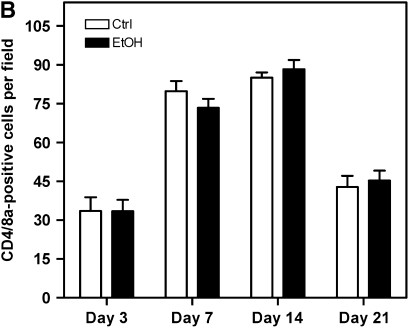
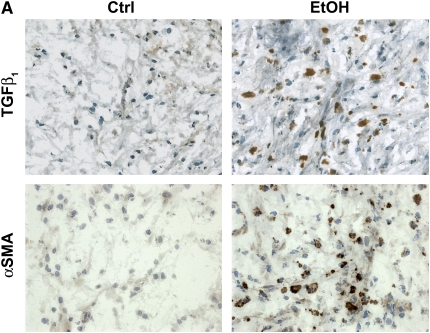
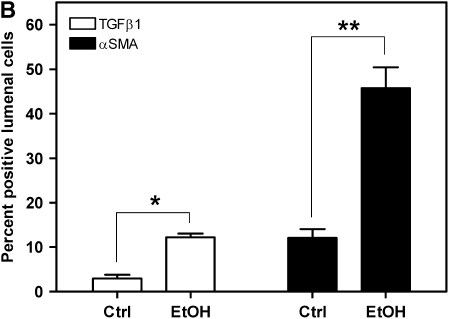
Measurements and main results: Although donor alcohol ingestion did not increase the number of antigen-presenting cells or infiltrating lymphocytes, it nevertheless increased allograft lumenal collagen content fourfold compared with allografts from control donors. In parallel, alcohol increased TGF-beta(1) and alpha-smooth muscle actin expression in allografts. Alcohol amplified airway disease even in isografts with minor alloimmune mismatches. In contrast, it did not cause any airway disease in isografts in a pure isogenic background, suggesting that a minimal alloimmune response is necessary to trigger alcohol-induced airway fibrosis.
Conclusions: Although alloimmune inflammation is required to initiate airway disease, alcohol primes the allograft for greater TGF-beta(1) expression, myofibroblast transdifferentiation, and fibrosis than by alloimmune inflammation alone. This has serious clinical implications, as many lung donors have underlying alcohol abuse that may prime the allograft recipient for subsequent OB.
Figures
References
-
- Boehler A, Kesten S, Weder W, Speich R. Bronchiolitis obliterans after lung transplantation: a review. Chest 1998;114:1411–1426. - PubMed
-
- Boehler A, Estenne M. Post-transplant bronchiolitis obliterans. Eur Respir J 2003;22:1007–1018. - PubMed
-
- Sharples LD, McNeil K, Stewart S, Wallwork J. Risk factors for bronchiolitis obliterans: a systematic review of recent publications. J Heart Lung Transplant 2002;21:271–281. - PubMed
-
- Scholma J, Slebos DJ, Boezen HM, van den Berg JW, van der Bij W, de Boer WJ, Koeter GH, Timens W, Kauffman HF, Postma DS. Eosinophilic granulocytes and interleukin-6 level in bronchoalveolar lavage fluid are associated with the development of obliterative bronchiolitis after lung transplantation. Am J Respir Crit Care Med 2000;162:2221–2225. - PubMed
-
- DiGiovine B, Lynch JP III, Martinez FJ, Flint A, Whyte RI, Iannettoni MD, Arenberg DA, Burdick MD, Glass MC, Wilke CA, et al. Bronchoalveolar lavage neutrophilia is associated with obliterative bronchiolitis after lung transplantation: role of IL-8. J Immunol 1996;157:4194–4202. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
