

Analysis of the role of retrotransposition in gene evolution in vertebrates
- PMID: 17718914
- PMCID: PMC2048973
- DOI: 10.1186/1471-2105-8-308
Analysis of the role of retrotransposition in gene evolution in vertebrates
Abstract
Background: The dynamics of gene evolution are influenced by several genomic processes. One such process is retrotransposition, where an mRNA transcript is reverse-transcribed and reintegrated into the genomic DNA.
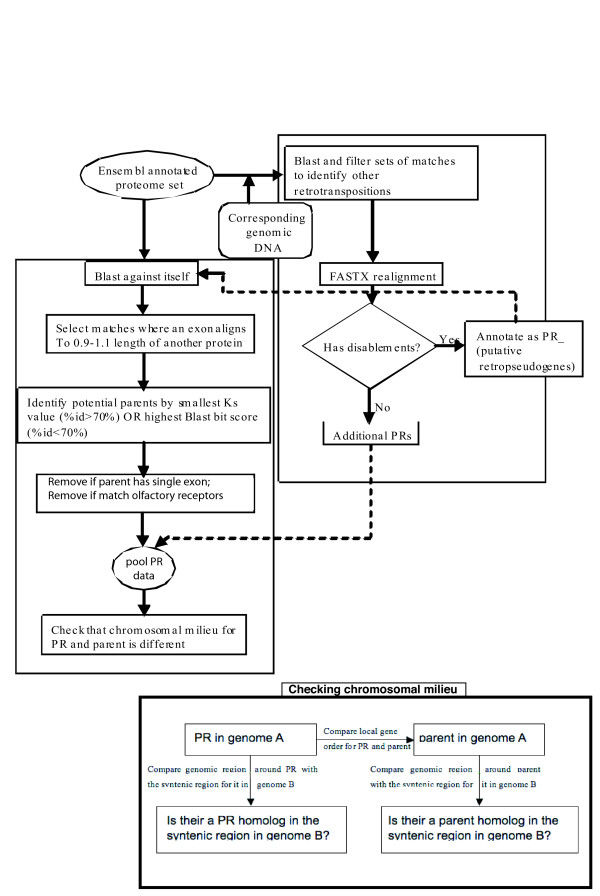
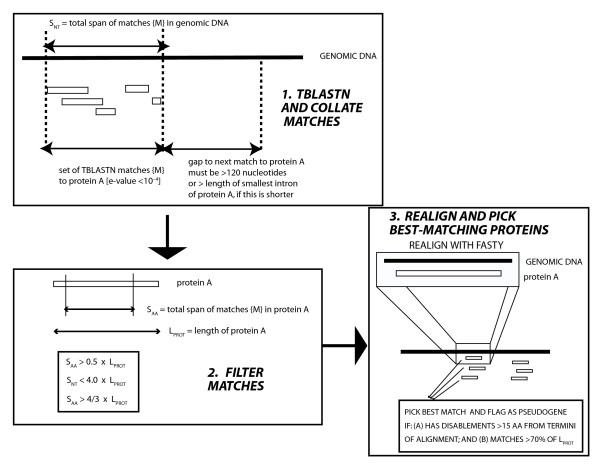
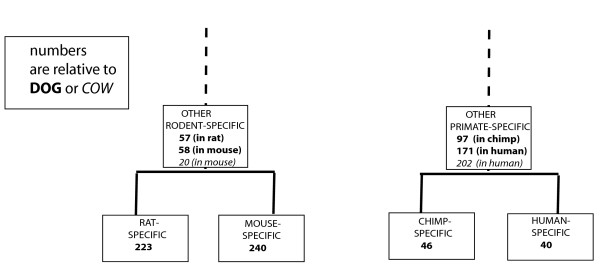
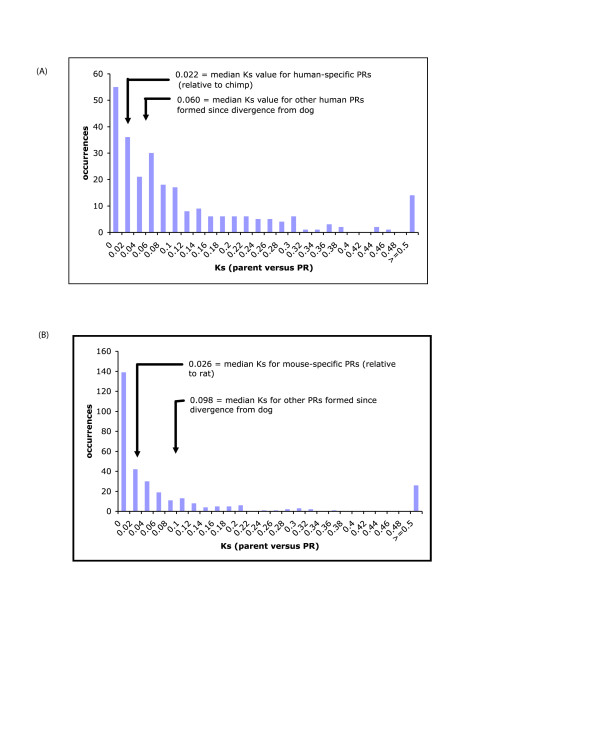
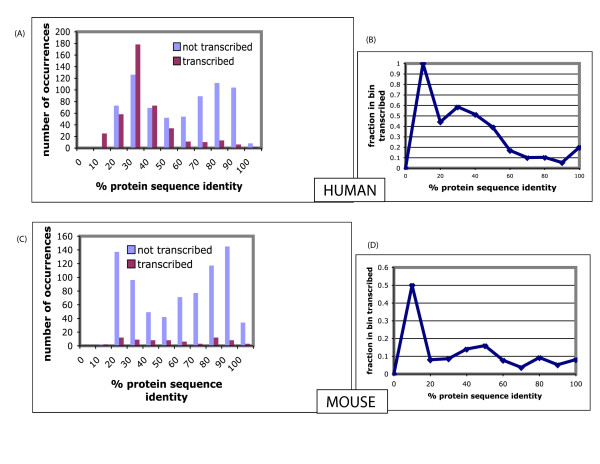
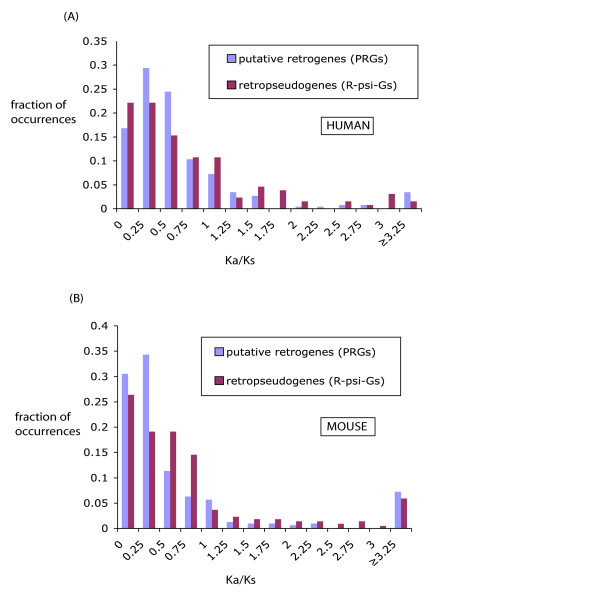
Results: We have surveyed eight vertebrate genomes (human, chimp, dog, cow, rat, mouse, chicken and the puffer-fish T. nigriviridis), for putatively retrotransposed copies of genes. To gain a complete picture of the role of retrotransposition, a robust strategy to identify putative retrogenes (PRs) was derived, in tandem with an adaptation of previous procedures to annotate processed pseudogenes, also called retropseudogenes (RpsiGs). Mammalian genomes are estimated to contain 400-800 PRs (corresponding to approximately 3% of genes), with fewer PRs and RpsiGs in the non-mammalian vertebrates. Focussing on human and mouse, we aged the PRs, analysed for evidence of transcription and selection pressures, and assigned functional categories. The PRs have significantly less transcription evidence mappable to them, are significantly less likely to arise from alternatively-spliced genes, and are statistically overrepresented for ribosomal-protein genes, when compared to the proteome in general. We find evidence for spurts of gene retrotransposition in human and mouse, since the lineage of either species split from the dog lineage, with >200 PRs formed in mouse since its divergence from rat. To examine for selection, we calculated: (i) Ka/Ks values (ratios of non-synonymous and synonymous substitutions in codons), and (ii) the significance of conservation of reading frames in PRs. We found >50 PRs in both human and mouse formed since divergence from dog, that are under pressure to maintain the integrity of their coding sequences. For different subsets of PRs formed at different stages of mammalian evolution, we find some evidence for non-neutral evolution, despite significantly less expression evidence for these sequences.
Conclusion: These results indicate that retrotranspositions are a significant source of novel coding sequences in mammalian gene evolution.
Figures
Similar articles
-
Assessing the genomic evidence for conserved transcribed pseudogenes under selection.BMC Genomics. 2009 Sep 15;10:435. doi: 10.1186/1471-2164-10-435. BMC Genomics. 2009. PMID: 19754956 Free PMC article.
-
Transcribed processed pseudogenes in the human genome: an intermediate form of expressed retrosequence lacking protein-coding ability.Nucleic Acids Res. 2005 Apr 28;33(8):2374-83. doi: 10.1093/nar/gki531. Print 2005. Nucleic Acids Res. 2005. PMID: 15860774 Free PMC article.
-
Frequent emergence and functional resurrection of processed pseudogenes in the human and mouse genomes.Gene. 2007 Mar 15;389(2):196-203. doi: 10.1016/j.gene.2006.11.007. Epub 2006 Nov 18. Gene. 2007. PMID: 17196768
-
L1 elements, processed pseudogenes and retrogenes in mammalian genomes.IUBMB Life. 2006 Dec;58(12):677-85. doi: 10.1080/15216540601034856. IUBMB Life. 2006. PMID: 17424906 Review.
-
Vertebrate pseudogenes.FEBS Lett. 2000 Feb 25;468(2-3):109-14. doi: 10.1016/s0014-5793(00)01199-6. FEBS Lett. 2000. PMID: 10692568 Review.
Cited by
-
Hidden treasures in unspliced EST data.Theory Biosci. 2012 May;131(1):49-57. doi: 10.1007/s12064-012-0151-6. Epub 2012 Apr 8. Theory Biosci. 2012. PMID: 22485013
-
Mitochondrial and nuclear localization of a novel pea thioredoxin: identification of its mitochondrial target proteins.Plant Physiol. 2009 Jun;150(2):646-57. doi: 10.1104/pp.109.138073. Epub 2009 Apr 10. Plant Physiol. 2009. PMID: 19363090 Free PMC article.
-
Genomic evidence for non-random endemic populations of decaying exons from mammalian genes.BMC Genomics. 2009 Jul 13;10:309. doi: 10.1186/1471-2164-10-309. BMC Genomics. 2009. PMID: 19594905 Free PMC article.
-
Comparative genomics, molecular evolution and computational modeling of ALDH1B1 and ALDH2.Chem Biol Interact. 2013 Feb 25;202(1-3):11-21. doi: 10.1016/j.cbi.2012.11.022. Epub 2012 Dec 13. Chem Biol Interact. 2013. PMID: 23247008 Free PMC article.
-
The Concept of Cancer Stem Cells: Elaborating on ALDH1B1 as an Emerging Marker of Cancer Progression.Life (Basel). 2023 Jan 9;13(1):197. doi: 10.3390/life13010197. Life (Basel). 2023. PMID: 36676146 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
