

Autocrine VEGF signaling is required for vascular homeostasis
- PMID: 17719546
- PMCID: PMC3010851
- DOI: 10.1016/j.cell.2007.06.054
Autocrine VEGF signaling is required for vascular homeostasis
Abstract
Vascular endothelial growth factor (VEGF) is essential for developmental and pathological angiogenesis. Here we show that in the absence of any pathological insult, autocrine VEGF is required for the homeostasis of blood vessels in the adult. Genetic deletion of vegf specifically in the endothelial lineage leads to progressive endothelial degeneration and sudden death in 55% of mutant mice by 25 weeks of age. The phenotype is manifested without detectable changes in the total levels of VEGF mRNA or protein, indicating that paracrine VEGF could not compensate for the absence of endothelial VEGF. Furthermore, wild-type, but not VEGF null, endothelial cells showed phosphorylation of VEGFR2 in the absence of exogenous VEGF. Activation of the receptor in wild-type cells was suppressed by small molecule antagonists but not by extracellular blockade of VEGF. These results reveal a cell-autonomous VEGF signaling pathway that holds significance for vascular homeostasis but is dispensable for the angiogenic cascade.
Figures
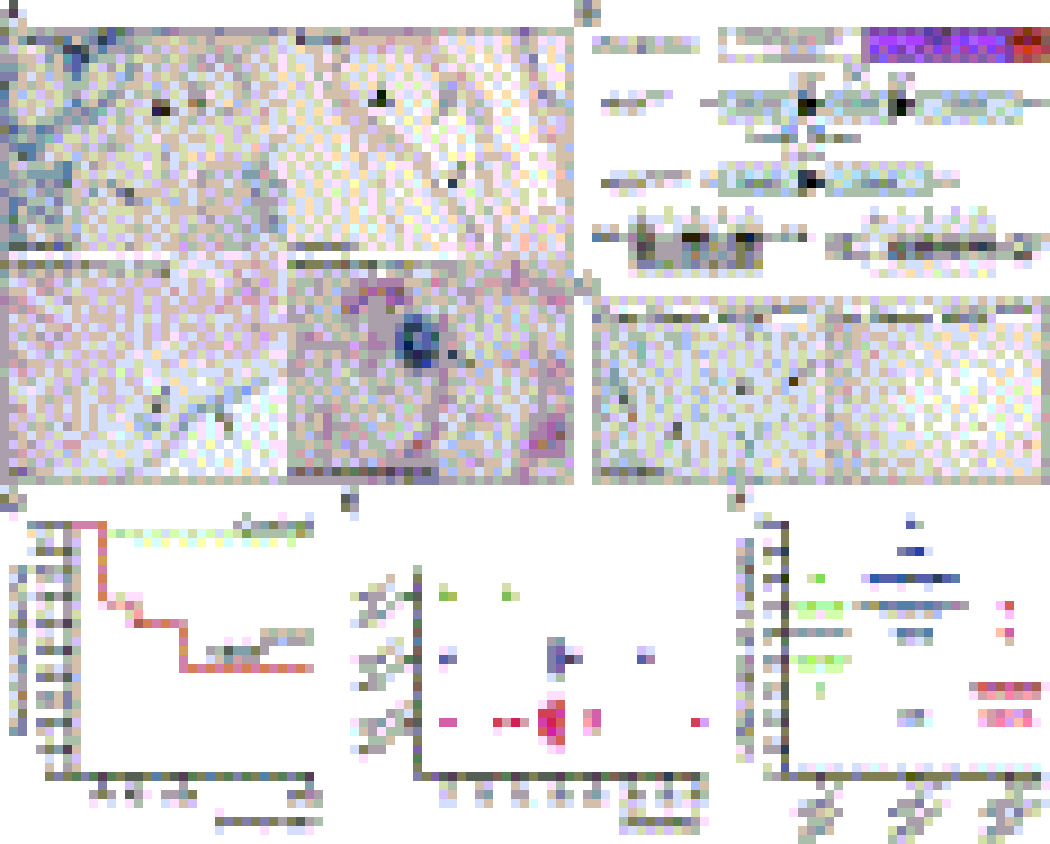

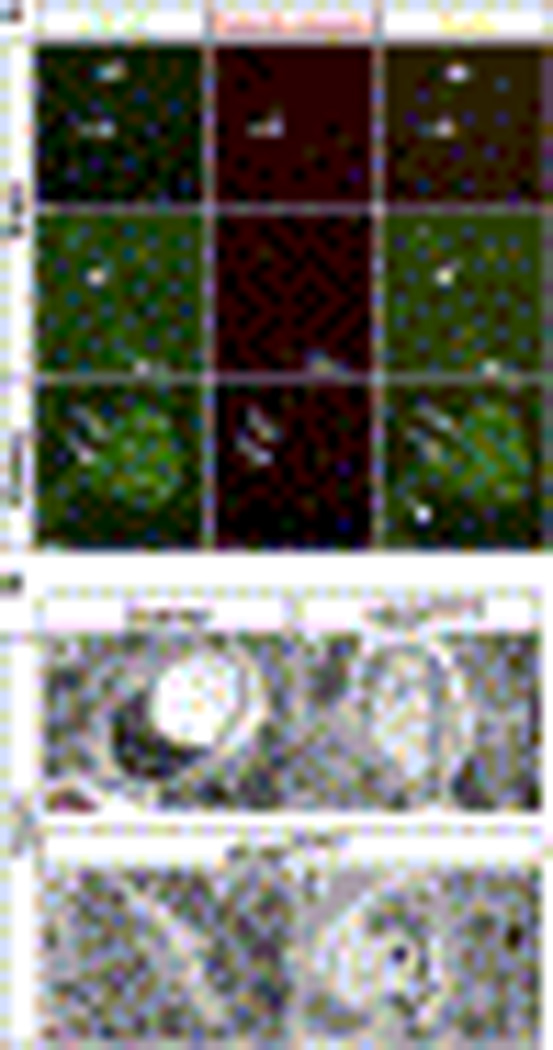

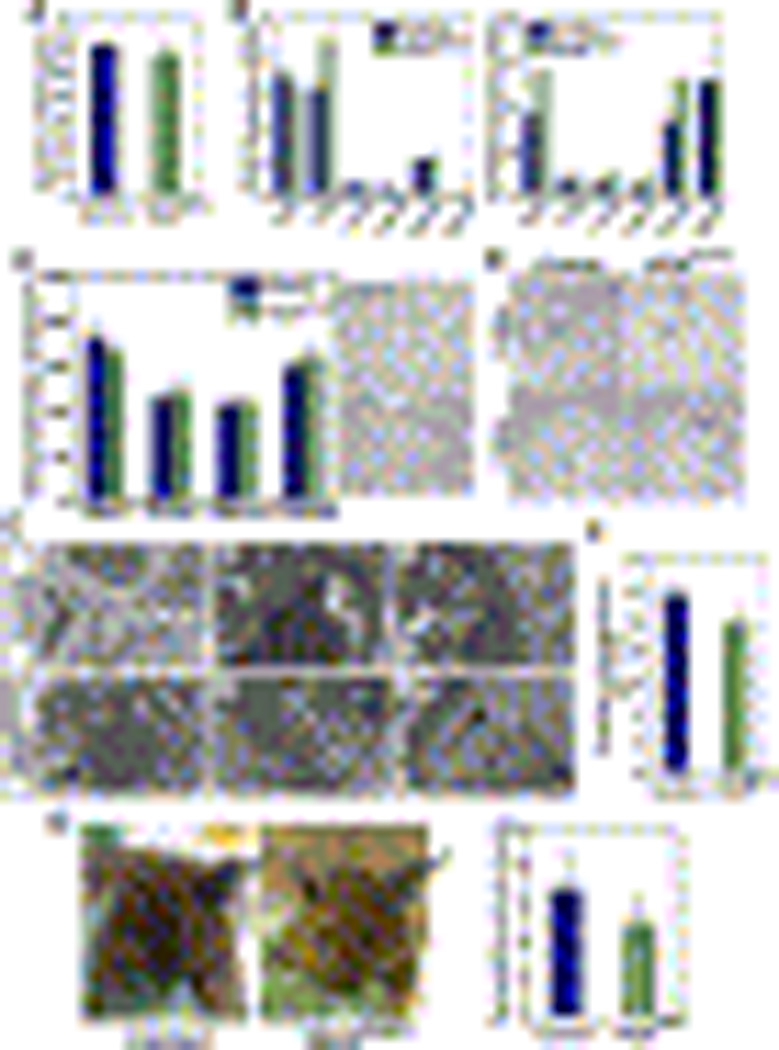
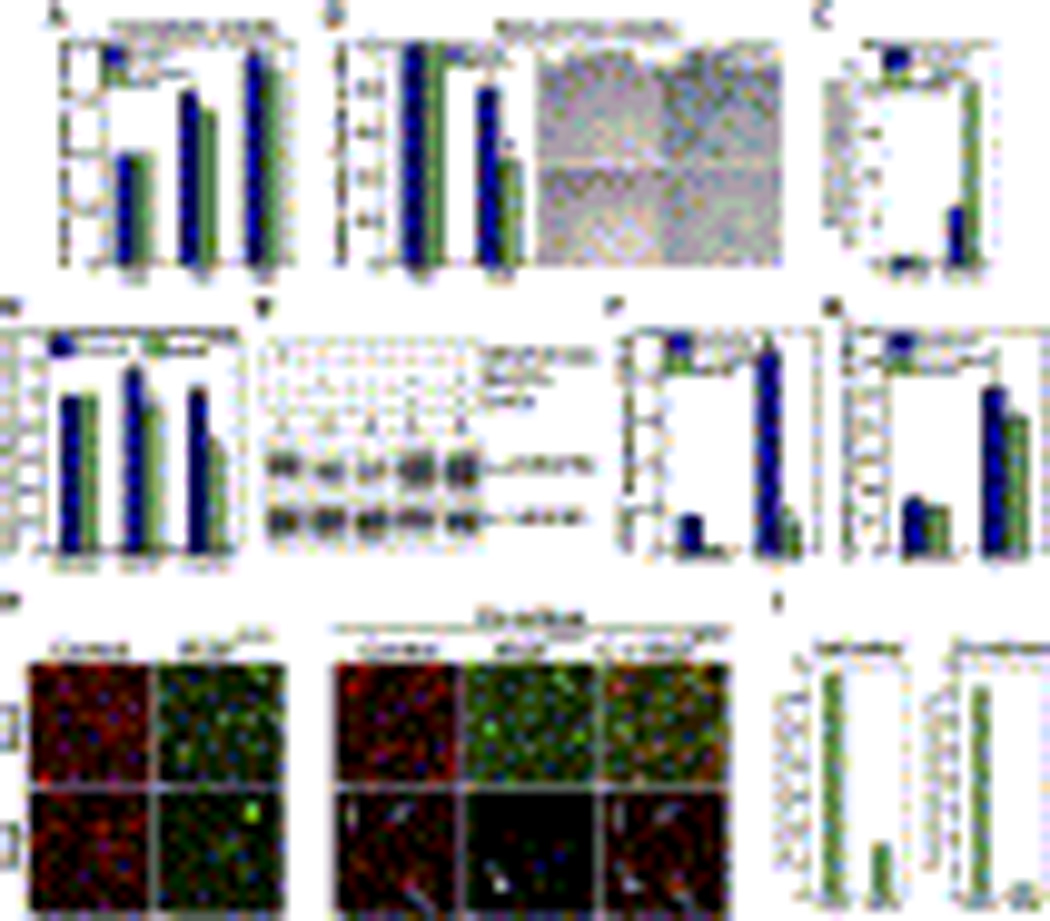
Comment in
-
The VEGF family, the inside story.Cell. 2007 Aug 24;130(4):591-2. doi: 10.1016/j.cell.2007.08.012. Cell. 2007. PMID: 17719536
References
-
- Alva JA, Zovein AC, Monvoisin A, Murphy T, Salazar A, Harvey NL, Carmeliet P, Iruela-Arispe ML. VE-Cadherin-Cre-recombinase transgenic mouse: a tool for lineage analysis and gene deletion in endothelial cells. Dev Dyn. 2006;235:759–767. - PubMed
-
- Bates D, Taylor GI, Minichiello J, Farlie P, Cichowitz A, Watson N, Klagsbrun M, Mamluk R, Newgreen DF. Neurovascular congruence results from a shared patterning mechanism that utilizes Semaphorin3A and Neuropilin-1. Dev Biol. 2003;255:77–98. - PubMed
-
- Blann AD. Assessment of endothelial dysfunction: focus on atherothrombotic disease. Pathophysiol Haemost Thromb. 2003a;33:256–261. - PubMed
-
- Blann AD. How a damaged blood vessel wall contibutes to thrombosis and hypertenasion. Pathophysiol Haemost Thromb. 2003b;33:445–448. - PubMed
-
- Bokel C, Schwabedissen A, Entchev E, Renaud O, Gonzalez-Gaitan M. Sara endosomes and the maintenance of Dpp signaling levels across mitosis. Science. 2006;314:1135–1139. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
