

Intrinsic circadian clock of the mammalian retina: importance for retinal processing of visual information
- PMID: 17719549
- PMCID: PMC2040024
- DOI: 10.1016/j.cell.2007.06.045
Intrinsic circadian clock of the mammalian retina: importance for retinal processing of visual information
Abstract
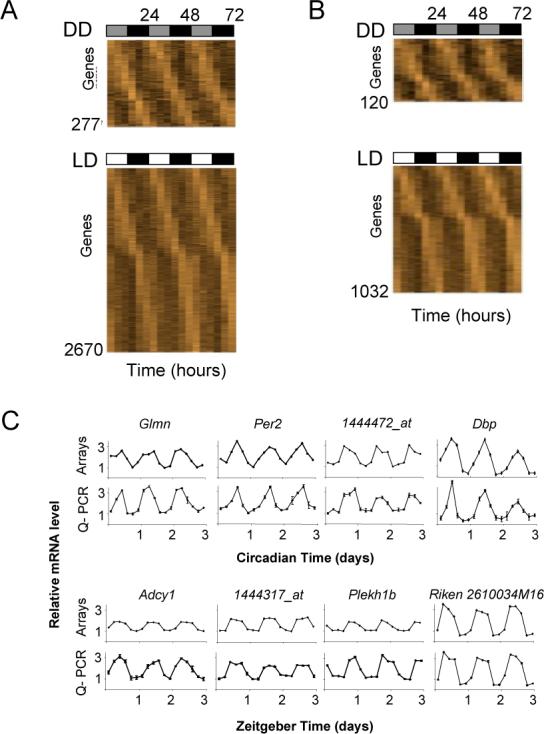
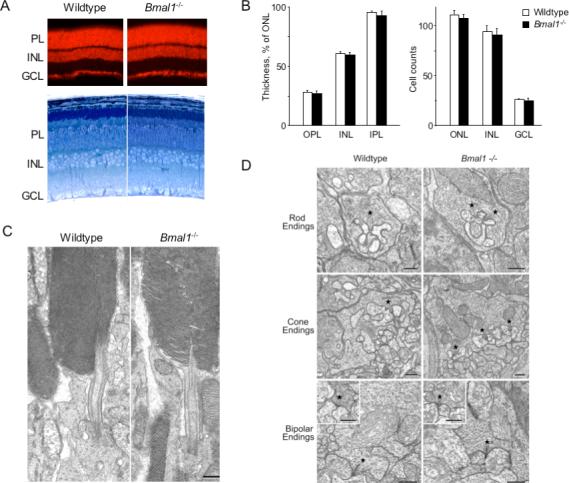
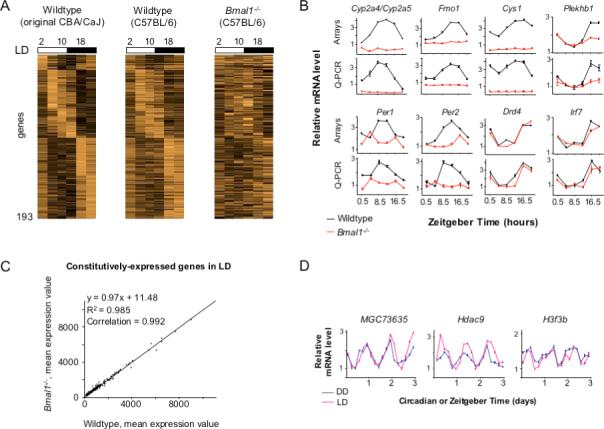
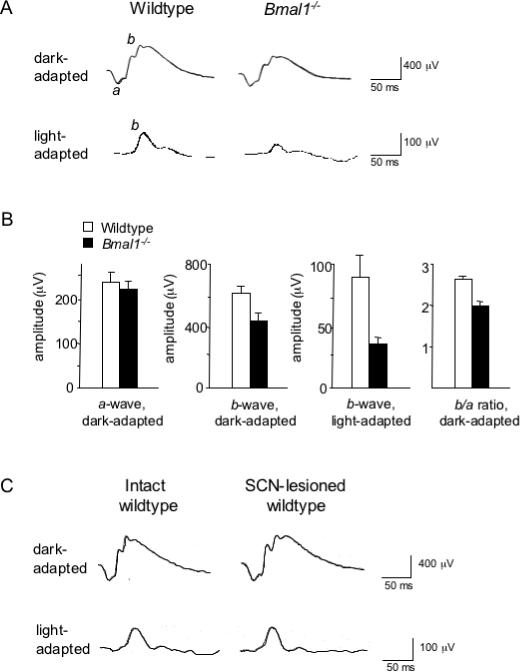
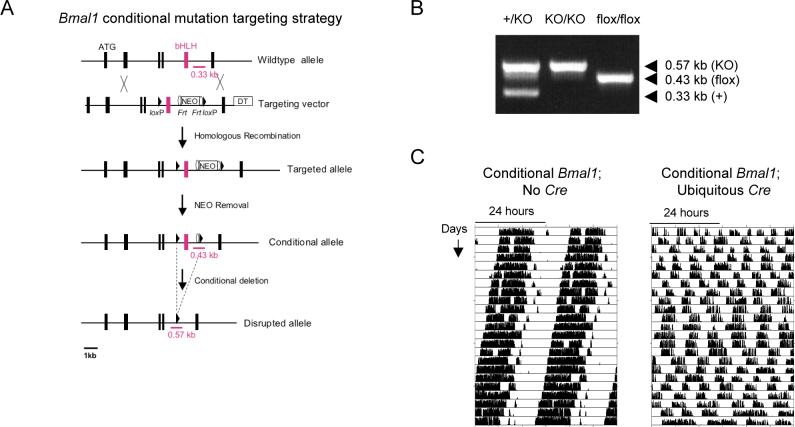
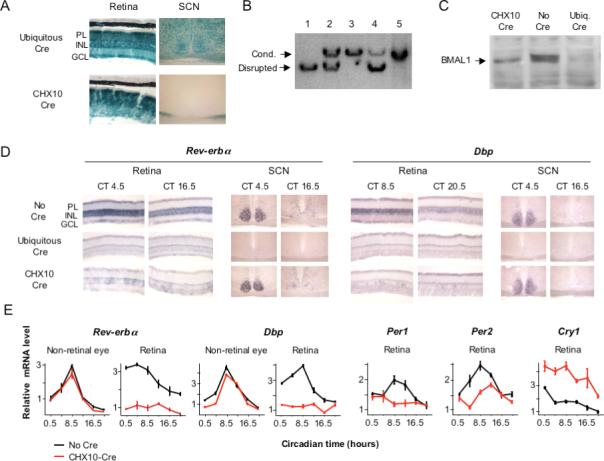
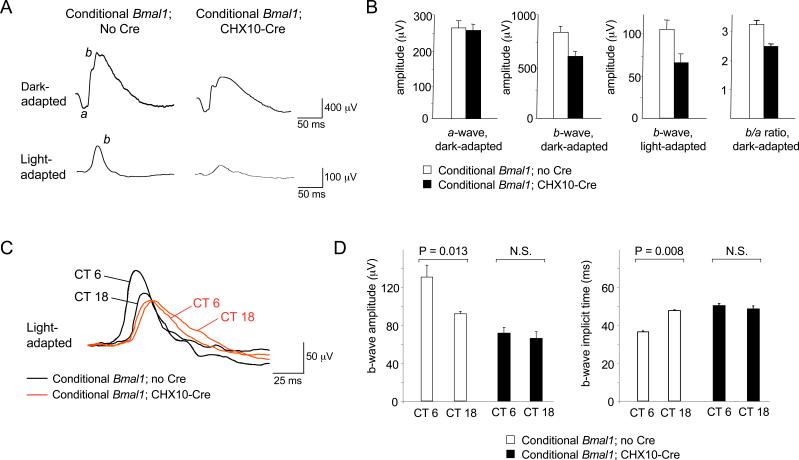
Circadian clocks are widely distributed in mammalian tissues, but little is known about the physiological functions of clocks outside the suprachiasmatic nucleus of the brain. The retina has an intrinsic circadian clock, but its importance for vision is unknown. Here we show that mice lacking Bmal1, a gene required for clock function, had abnormal retinal transcriptional responses to light and defective inner retinal electrical responses to light, but normal photoreceptor responses to light and retinas that appeared structurally normal by light and electron microscopy. We generated mice with a retina-specific genetic deletion of Bmal1, and they had defects of retinal visual physiology essentially identical to those of mice lacking Bmal1 in all tissues and lacked a circadian rhythm of inner retinal electrical responses to light. Our findings indicate that the intrinsic circadian clock of the retina regulates retinal visual processing in vivo.
Figures
References
-
- Akhtar RA, Reddy AB, Maywood ES, Clayton JD, King VM, Smith AG, Gant TW, Hastings MH, Kyriacou CP. Circadian cycling of the mouse liver transcriptome, as revealed by cDNA microarray, is driven by the suprachiasmatic nucleus. Curr. Biol. 2002;12:540–550. - PubMed
-
- Bailey MJ, Beremand PD, Hammer R, Reidel E, Thomas TL, Cassone VM. Transcriptional profiling of circadian patterns of mRNA expression in the chick retina. J. Biol. Chem. 2004;279:52247–52254. - PubMed
-
- Barnard AR, Hattar S, Hankins MW, Lucas RJ. Melanopsin regulates visual processing in the mouse retina. Curr. Biol. 2006;16:389–395. - PubMed
-
- Balsalobre A, Damiola F, Schibler U. A serum shock induces circadian gene expression in cultured Rat-1 fibroblasts. Cell. 1998;93:929–937. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
