

Inhibition of staphylococcal biofilm formation by nitrite
- PMID: 17720780
- PMCID: PMC2168742
- DOI: 10.1128/JB.00598-07
Inhibition of staphylococcal biofilm formation by nitrite
Abstract
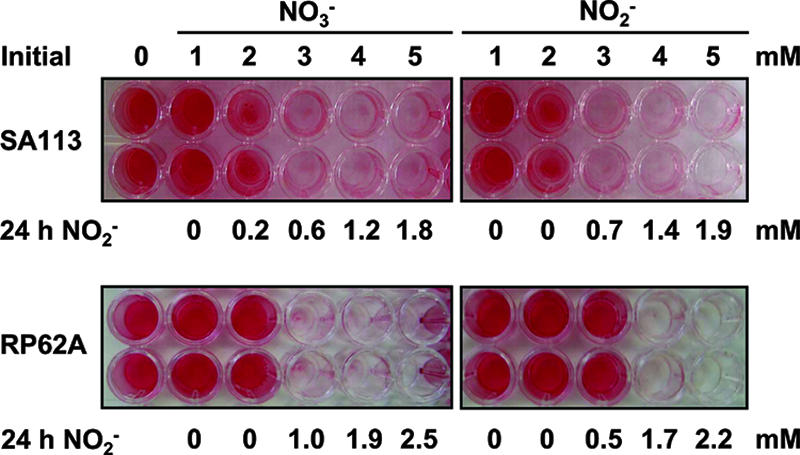
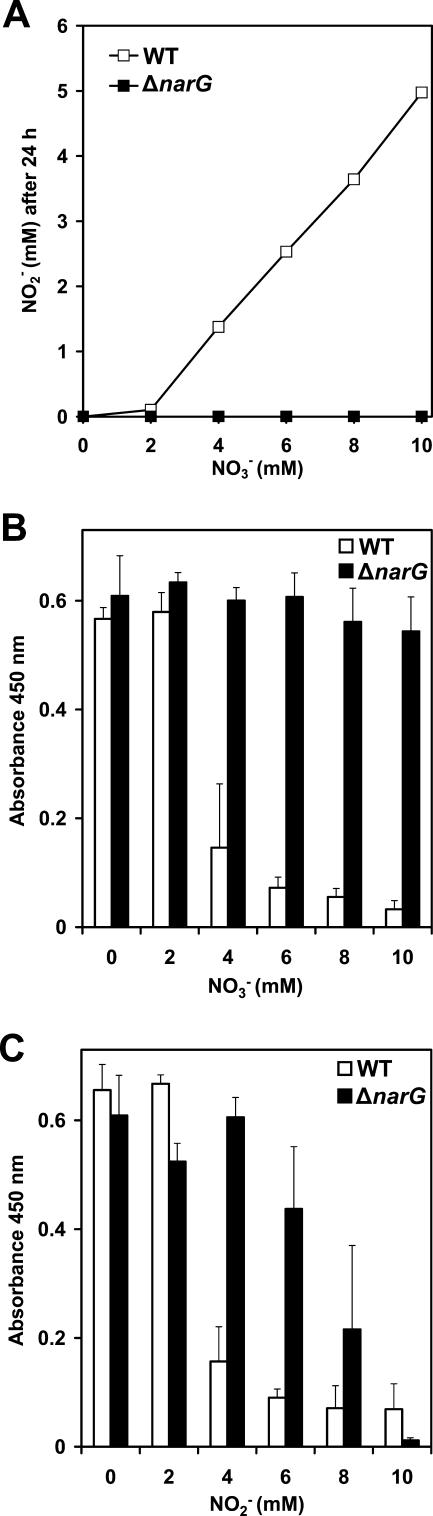
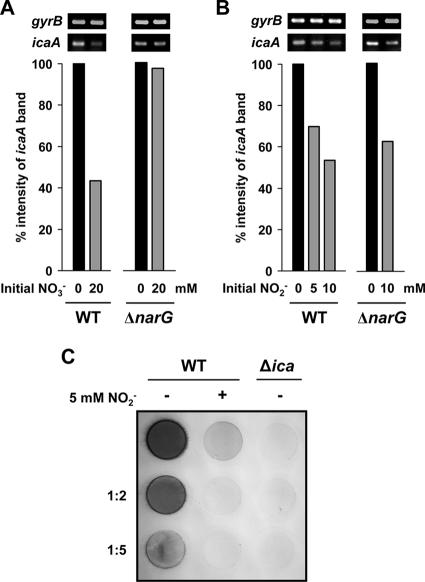
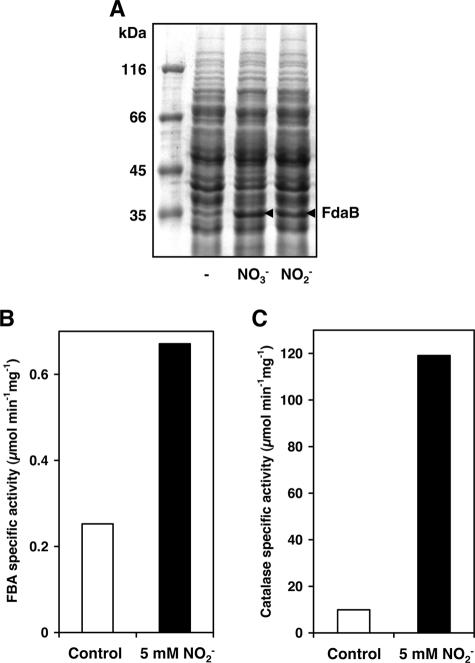
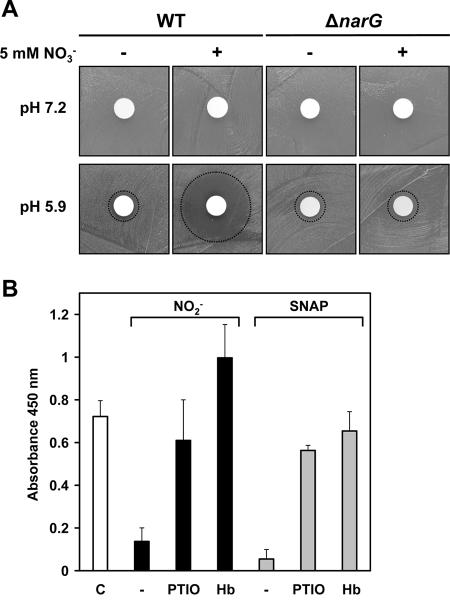
Several environmental stresses have been demonstrated to increase polysaccharide intercellular adhesin (PIA) synthesis and biofilm formation by the human pathogens Staphylococcus aureus and Staphylococcus epidermidis. In this study we characterized an adaptive response of S. aureus SA113 to nitrite-induced stress and show that it involves concomitant impairment of PIA synthesis and biofilm formation. Transcriptional analysis provided evidence that nitrite, either as the endogenous product of respiratory nitrate reduction or after external addition, causes repression of the icaADBC gene cluster, mediated likely by IcaR. Comparative microarray analysis revealed a global change in gene expression during growth in the presence of 5 mM sodium nitrite and indicated a response to oxidative and nitrosative stress. Many nitrite-induced genes are involved in DNA repair, detoxification of reactive oxygen and nitrogen species, and iron homeostasis. Moreover, preformed biofilms could be eradicated by the addition of nitrite, likely the result of the formation of toxic acidified nitrite derivatives. Nitrite-mediated inhibition of S. aureus biofilm formation was abrogated by the addition of nitric oxide (NO) scavengers, suggesting that NO is directly or indirectly involved. Nitrite also repressed biofilm formation of S. epidermidis RP62A.
Figures
Similar articles
-
Polysaccharide intercellular adhesin in biofilm: structural and regulatory aspects.Front Cell Infect Microbiol. 2015 Feb 10;5:7. doi: 10.3389/fcimb.2015.00007. eCollection 2015. Front Cell Infect Microbiol. 2015. PMID: 25713785 Free PMC article. Review.
-
Spontaneous switch to PIA-independent biofilm formation in an ica-positive Staphylococcus epidermidis isolate.Int J Med Microbiol. 2007 Apr;297(2):117-22. doi: 10.1016/j.ijmm.2006.12.001. Epub 2007 Feb 9. Int J Med Microbiol. 2007. PMID: 17292669
-
Anti-biofilm activity of sub-inhibitory povidone-iodine concentrations against Staphylococcus epidermidis and Staphylococcus aureus.J Orthop Res. 2010 Sep;28(9):1252-6. doi: 10.1002/jor.21110. J Orthop Res. 2010. PMID: 20187117
-
The staphylococcal respiratory response regulator SrrAB induces ica gene transcription and polysaccharide intercellular adhesin expression, protecting Staphylococcus aureus from neutrophil killing under anaerobic growth conditions.Mol Microbiol. 2007 Sep;65(5):1276-87. doi: 10.1111/j.1365-2958.2007.05863.x. Mol Microbiol. 2007. PMID: 17697253
-
Mechanisms of biofilm formation in Staphylococcus epidermidis and Staphylococcus aureus: functional molecules, regulatory circuits, and adaptive responses.Int J Med Microbiol. 2004 Sep;294(2-3):203-12. doi: 10.1016/j.ijmm.2004.06.015. Int J Med Microbiol. 2004. PMID: 15493831 Review.
Cited by
-
Origin and Impact of Nitric Oxide in Pseudomonas aeruginosa Biofilms.J Bacteriol. 2016 Jan 1;198(1):55-65. doi: 10.1128/JB.00371-15. J Bacteriol. 2016. PMID: 26260455 Free PMC article. Review.
-
Oxidation-sensing regulator AbfR regulates oxidative stress responses, bacterial aggregation, and biofilm formation in Staphylococcus epidermidis.J Biol Chem. 2013 Feb 8;288(6):3739-52. doi: 10.1074/jbc.M112.426205. Epub 2012 Dec 27. J Biol Chem. 2013. PMID: 23271738 Free PMC article.
-
Increased Antibacterial and Antibiofilm Properties of Silver Nanoparticles Using Silver Fluoride as Precursor.Molecules. 2020 Jul 31;25(15):3494. doi: 10.3390/molecules25153494. Molecules. 2020. PMID: 32751978 Free PMC article.
-
Transcriptional profiling identifies the metabolic phenotype of gonococcal biofilms.Infect Immun. 2009 Sep;77(9):3522-32. doi: 10.1128/IAI.00036-09. Epub 2009 Jun 15. Infect Immun. 2009. PMID: 19528210 Free PMC article.
-
Broad impact of extracellular DNA on biofilm formation by clinically isolated Methicillin-resistant and -sensitive strains of Staphylococcus aureus.Sci Rep. 2018 Feb 2;8(1):2254. doi: 10.1038/s41598-018-20485-z. Sci Rep. 2018. PMID: 29396526 Free PMC article.
References
-
- Anderson, K. L., C. Roberts, T. Disz, V. Vonstein, K. Hwang, R. Overbeek, P. D. Olson, S. J. Projan, and P. M. Dunman. 2006. Characterization of the Staphylococcus aureus heat shock, cold shock, stringent, and SOS responses and their effects on log-phase mRNA turnover. J. Bacteriol. 188:6739-6756. - PMC - PubMed
-
- Augustin, J., and F. Götz. 1990. Transformation of Staphylococcus epidermidis and other staphylococcal species with plasmid DNA by electroporation. FEMS Microbiol. Lett. 54:203-207. - PubMed
-
- Beers, R. F., Jr., and I. W. Sizer. 1952. A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase. J. Biol. Chem. 195:133-140. - PubMed
-
- Biswas, R., L. Voggu, U. K. Simon, P. Hentschel, G. Thumm, and F. Götz. 2006. Activity of the major staphylococcal autolysin Atl. FEMS Microbiol. Lett. 259:260-268. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
