

Role of dendritic cells in differential susceptibility to viral demyelinating disease
- PMID: 17722981
- PMCID: PMC1950949
- DOI: 10.1371/journal.ppat.0030124
Role of dendritic cells in differential susceptibility to viral demyelinating disease
Abstract
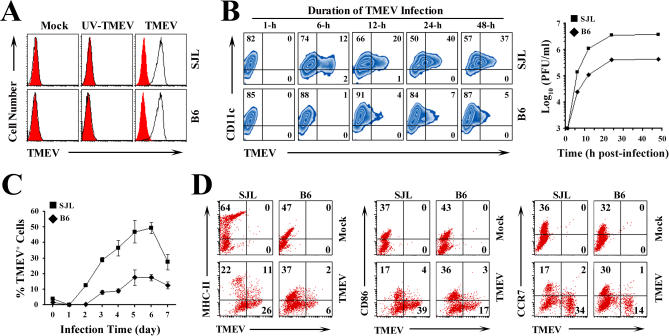
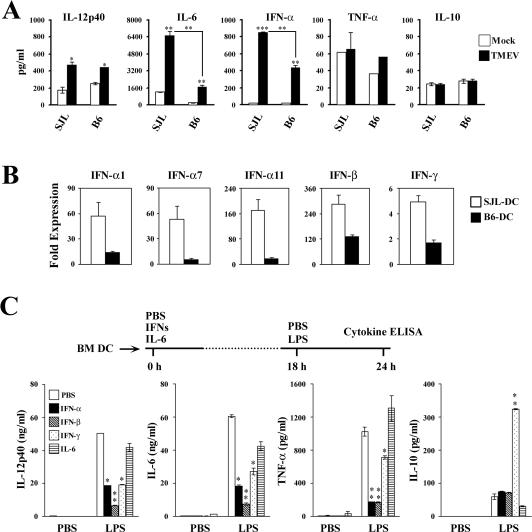
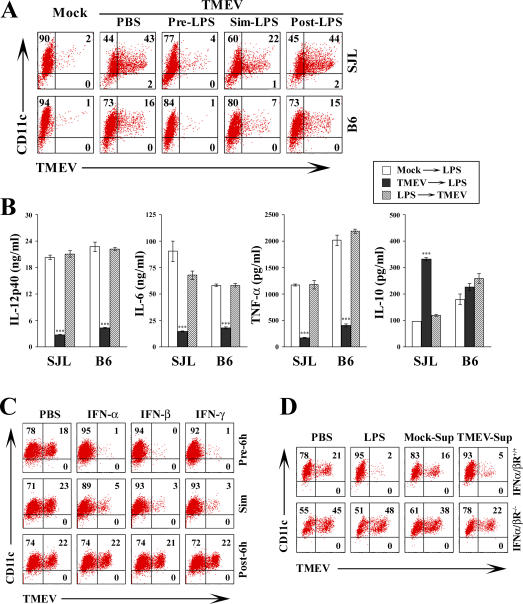
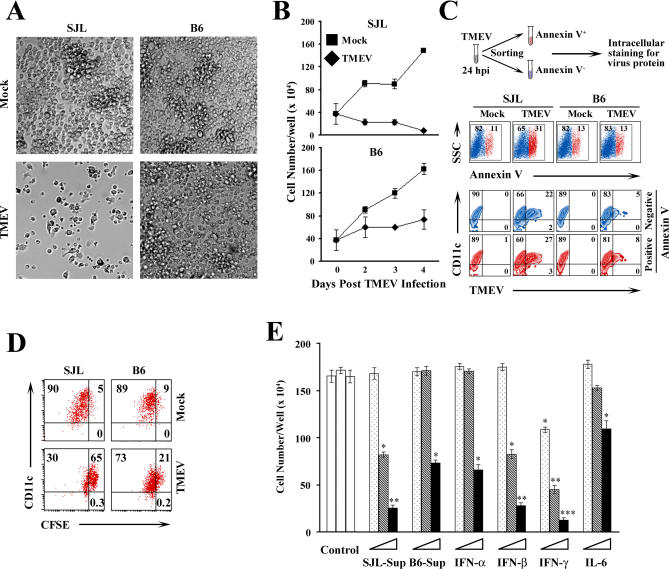
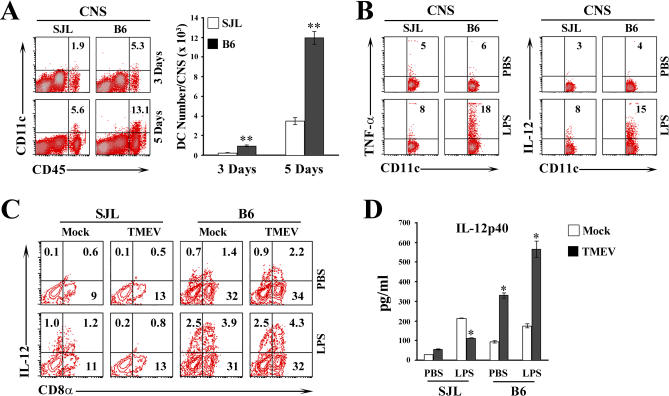
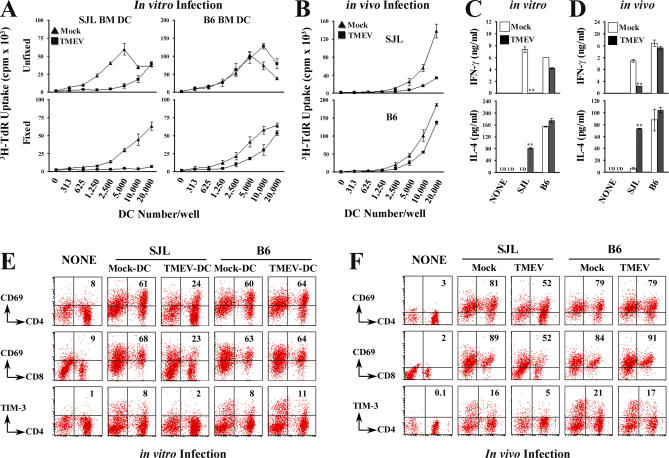
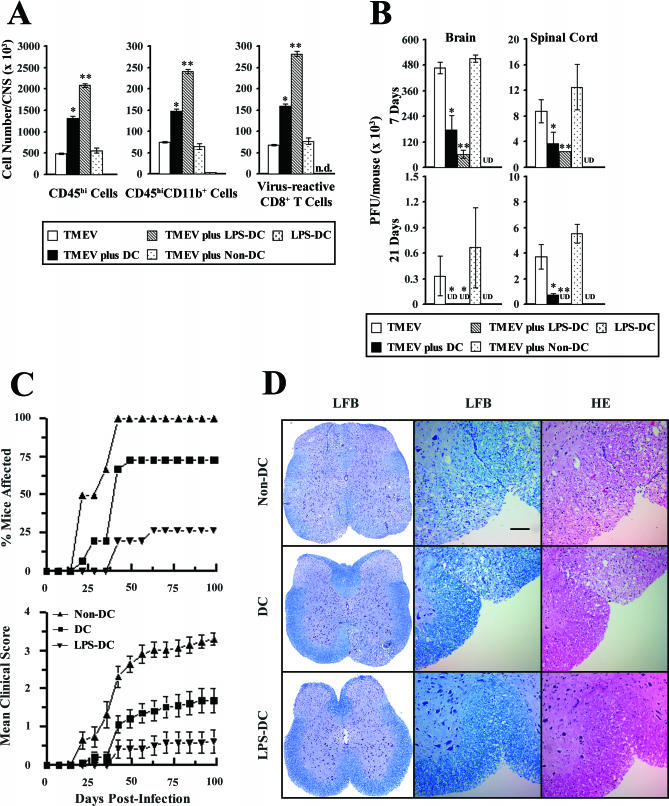
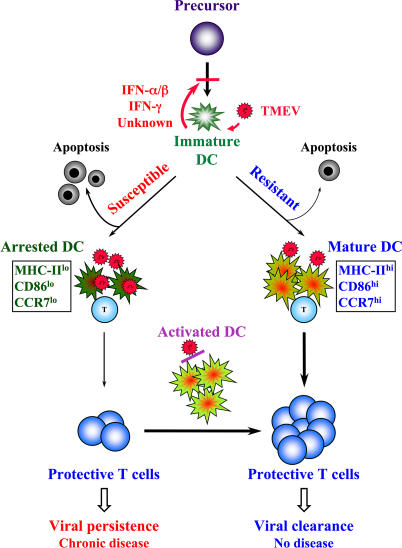
Although persistent viral diseases are a global health concern, the mechanisms of differential susceptibility to such infections among individuals are unknown. Here, we report that differential interactions between dendritic cells (DCs) and virus are critical in determining resistance versus susceptibility in the Theiler murine encephalomyelitis virus-induced demyelinating disease model of multiple sclerosis. This virus induces a chronic demyelinating disease in susceptible mice, whereas the virus is completely cleared in resistant strains of mice. DCs from susceptible mice are more permissive to viral infection, resulting in severe deficiencies in development, expansion, and function, in contrast to DCs from resistant mice. Although protective prior to viral infection, higher levels of type I interferons (IFNs) and IFN-gamma produced by virus-infected DCs from susceptible mice further contribute to the differential inhibition of DC development and function. An increased DC number and/or acquired resistance of DCs to viral infection render susceptible mice resistant to viral persistence and disease progression. Thus, the differential permissiveness of DCs to infectious agents and its subsequent functional and developmental deficiencies determine the outcome of infection- associated diseases. Therefore, arming DCs against viral infection-induced functional decline may provide a useful intervention for chronic infection-associated diseases.
Conflict of interest statement
Figures
References
-
- Hill AV. The immunogenetics of human infectious diseases. Annu Rev Immunol. 1998;16:593–617. - PubMed
-
- Beutler B, Jiang Z, Georgel P, Crozat K, Croker B, et al. Genetic analysis of host resistance: Toll-like receptor signaling and immunity at large. Annu Rev Immunol. 2006;24:353–389. - PubMed
-
- Granucci F, Foti M, Ricciardi-Castagnoli P. Dendritic cell biology. Adv Immunol. 2005;88:193–233. - PubMed
-
- Dal Canto MC, Kim B.S, Miller S.D, Melvold R.W. Theiler's murine encephalomyelitis virus (TMEV)-induced demyelination: A model for human multiple clerosis. Methods. 1996;10:453–461. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
