

Early endocardial morphogenesis requires Scl/Tal1
- PMID: 17722983
- PMCID: PMC1950955
- DOI: 10.1371/journal.pgen.0030140
Early endocardial morphogenesis requires Scl/Tal1
Abstract
The primitive heart tube is composed of an outer myocardial and an inner endocardial layer that will give rise to the cardiac valves and septa. Specification and differentiation of these two cell layers are among the earliest events in heart development, but the embryonic origins and genetic regulation of early endocardial development remain largely undefined. We have analyzed early endocardial development in the zebrafish using time-lapse confocal microscopy and show that the endocardium seems to originate from a region in the lateral plate mesoderm that will give rise to hematopoietic cells of the primitive myeloid lineage. Endocardial precursors appear to rapidly migrate to the site of heart tube formation, where they arrive prior to the bilateral myocardial primordia. Analysis of a newly discovered zebrafish Scl/Tal1 mutant showed an additional and previously undescribed role of this transcription factor during the development of the endocardium. In Scl/Tal1 mutant embryos, endocardial precursors are specified, but migration is severely defective and endocardial cells aggregate at the ventricular pole of the heart. We further show that the initial fusion of the bilateral myocardial precursor populations occurs independently of the endocardium and tal1 function. Our results suggest early separation of the two components of the primitive heart tube and imply Scl/Tal1 as an indispensable component of the molecular hierarchy that controls endocardium morphogenesis.
Conflict of interest statement
Competing interests. The authors have declared that no competing interests exist.
Figures
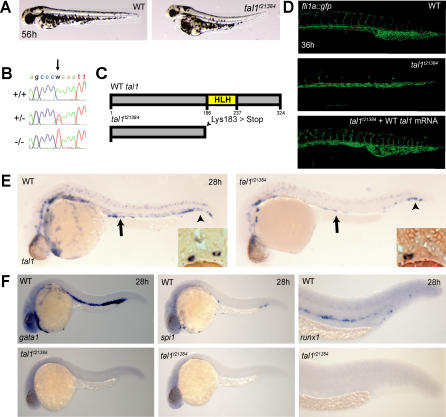

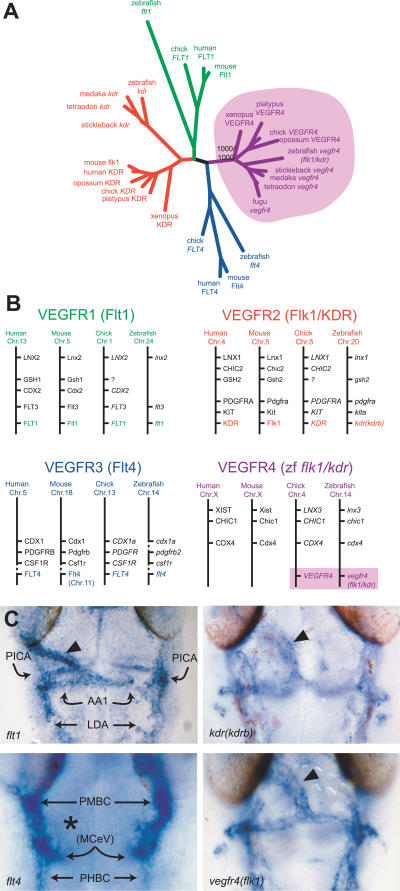

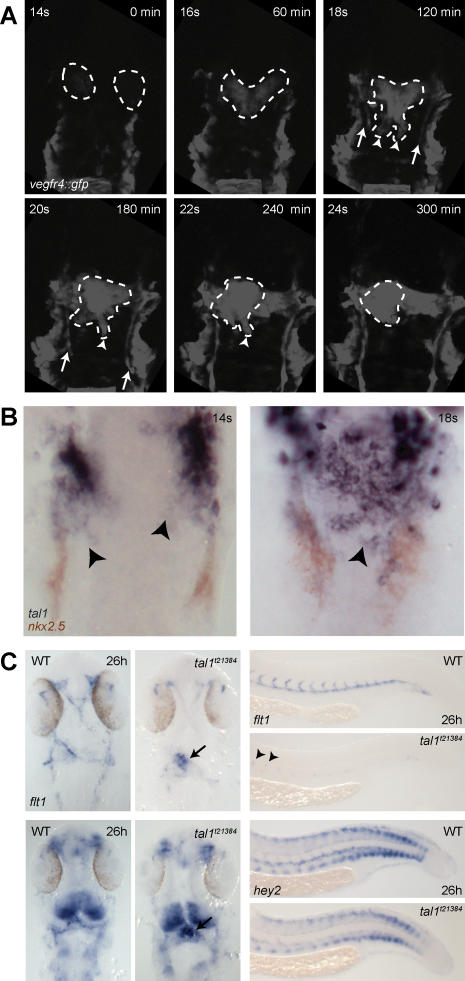

References
-
- Lough J, Sugi Y. Endoderm and heart development. Dev Dyn. 2000;217:327–342. - PubMed
-
- Kattman SJ, Huber TL, Keller GM. Multipotent flk-1(+) cardiovascular progenitor cells give rise to the cardiomyocyte, endothelial, and vascular smooth muscle lineages. Dev Cell. 2006;11:723–732. - PubMed
-
- Schultheiss TM, Burch JB, Lassar AB. A role for bone morphogenetic proteins in the induction of cardiac myogenesis. Genes Dev. 1997;11:451–462. - PubMed
-
- Nemer G, Nemer M. Cooperative interaction between GATA5 and NF-ATc regulates endothelial-endocardial differentiation of cardiogenic cells. Development. 2002;129:4045–4055. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
