

Identification of a region on hypoxia-inducible-factor prolyl 4-hydroxylases that determines their specificity for the oxygen degradation domains
- PMID: 17725546
- PMCID: PMC2267344
- DOI: 10.1042/BJ20071052
Identification of a region on hypoxia-inducible-factor prolyl 4-hydroxylases that determines their specificity for the oxygen degradation domains
Abstract
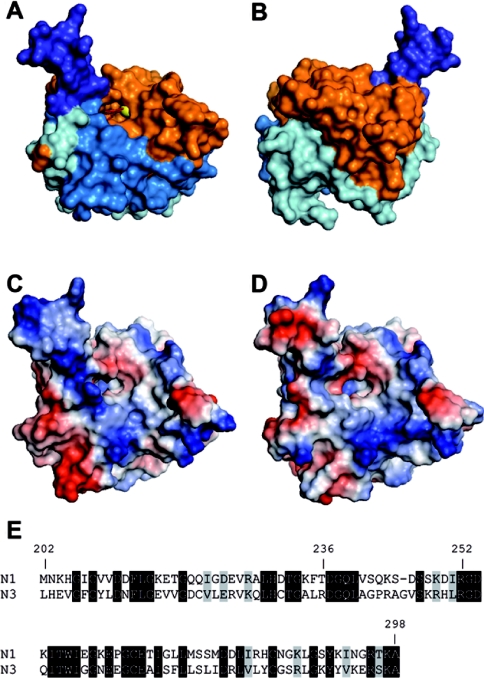
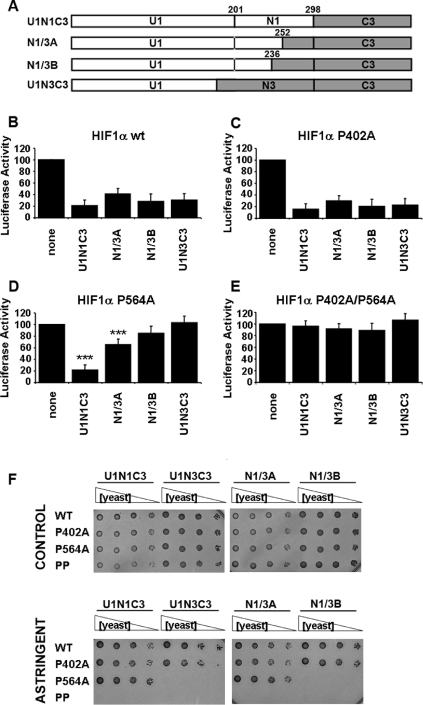
HIFs [hypoxia-inducible (transcription) factors] are essential for the induction of an adaptive gene expression programme under low oxygen partial pressure. The activity of these transcription factors is mainly determined by the stability of the HIFalpha subunit, which is regulated, in an oxygen-dependent manner, by a family of three prolyl 4-hydroxylases [EGLN1-EGLN3 (EGL nine homologues 1-3)]. HIFalpha contains two, N- and C-terminal, independent ODDs (oxygen-dependent degradation domains), namely NODD and CODD, that, upon hydroxylation by the EGLNs, target HIFalpha for proteasomal degradation. In vitro studies indicate that each EGLN shows a differential preference for ODDs, However, the sequence determinants for such specificity are unknown. In the present study we showed that whereas EGLN1 and EGLN2 acted upon any of these ODDs to regulate HIF1alpha protein levels and activity in vivo, EGLN3 only acted on the CODD. With the aim of identifying the region within EGLNs responsible for their differential substrate preference, we investigated the activity and binding pattern of different EGLN deletions and chimaeric constructs generated by domain swapping between EGLN1 and EGLN3. These studies revealed a region of 97 residues that was sufficient to confer the characteristic substrate binding observed for each EGLN. Within this region, we identified the minimal sequence (EGLN1 residues 236-252) involved in substrate discrimination. Importantly, mapping of these sequences on the EGLN1 tertiary structure indicates that substrate specificity is determined by a region relatively remote from the catalytic site.
Figures




Comment in
-
Enzyme substrate recognition in oxygen sensing: how the HIF trap snaps.Biochem J. 2007 Dec 1;408(2):e5-6. doi: 10.1042/BJ20071306. Biochem J. 2007. PMID: 17990984 Free PMC article. Review.
References
-
- Wenger R. H., Stiehl D. P., Camenisch G. Integration of oxygen signaling at the consensus HRE. Sci. STKE. 2005;306:re12. - PubMed
-
- Schofield C. J., Ratcliffe P. J. Oxygen sensing by HIF hydroxylases. Nat. Rev. Mol. Cell Biol. 2004;5:343–354. - PubMed
-
- Ivan M., Kondo K., Yang H., Kim W., Valiando J., Ohh M., Salic A., Asara J. M., Lane W. S., Kaelin W. G., Jr HIFα targeted for VHL-mediated destruction by proline hydroxylation: implications for O2 sensing. Science. 2001;292:464–468. - PubMed
-
- Jaakkola P., Mole D. R., Tian Y. M., Wilson M. I., Gielbert J., Gaskell S. J., Kriegsheim A., Hebestreit H. F., Mukherji M., Schofield C. J., et al. Targeting of HIF-α to the von Hippel–Lindau ubiquitylation complex by O2-regulated prolyl hydroxylation. Science. 2001;292:468–472. - PubMed
-
- Epstein A. C., Gleadle J. M., McNeill L. A., Hewitson K. S., O'Rourke J., Mole D. R., Mukherji M., Metzen E., Wilson M. I., Dhanda A., et al. C. elegans EGL-9 and mammalian homologs define a family of dioxygenases that regulate HIF by prolyl hydroxylation. Cell. 2001;107:43–54. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources