

Phosphorylation of synapsin domain A is required for post-tetanic potentiation
- PMID: 17726061
- PMCID: PMC3016615
- DOI: 10.1242/jcs.012005
Phosphorylation of synapsin domain A is required for post-tetanic potentiation
Erratum in
- J Cell Sci. 2007 Sep 15;120(Pt 18):3321
Abstract
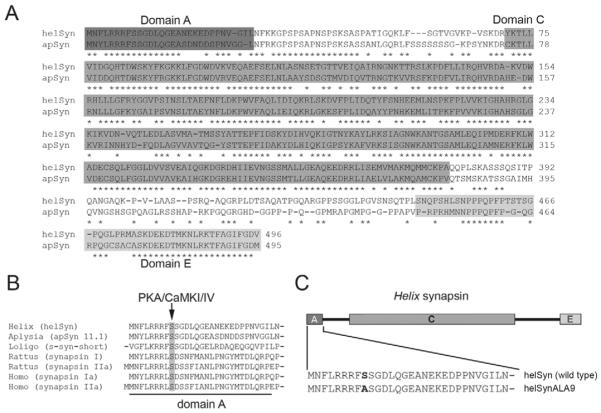
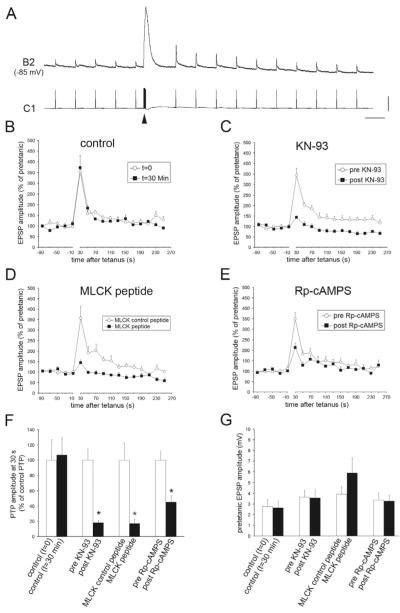
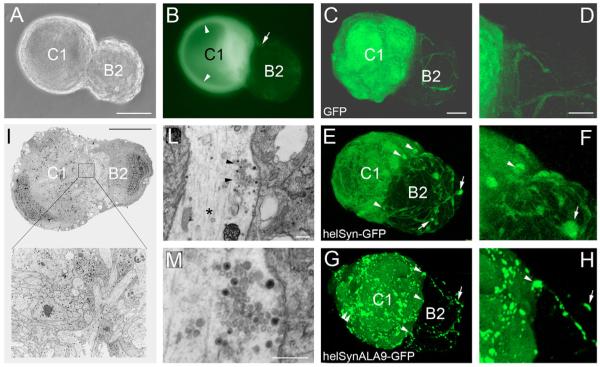
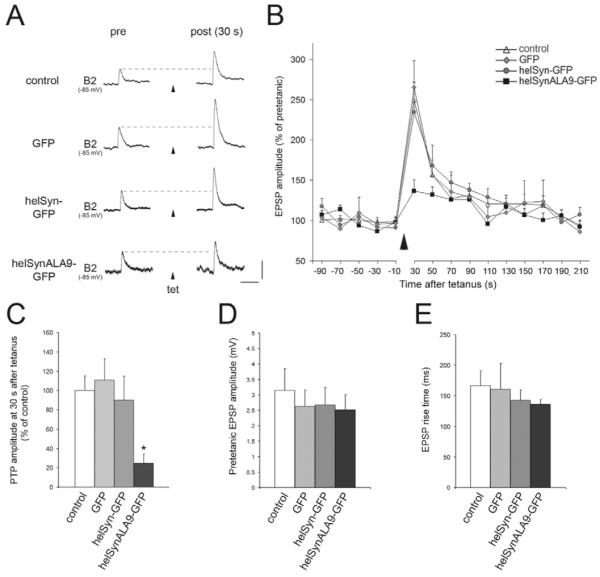
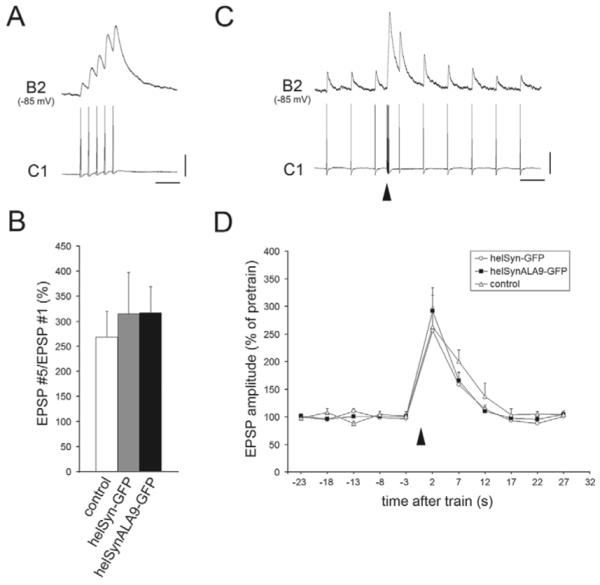
Post-tetanic potentiation (PTP) is a form of homosynaptic plasticity important for information processing and short-term memory in the nervous system. The synapsins, a family of synaptic vesicle (SV)-associated phosphoproteins, have been implicated in PTP. Although several synapsin functions are known to be regulated by phosphorylation by multiple protein kinases, the role of individual phosphorylation sites in synaptic plasticity is poorly understood. All the synapsins share a phosphorylation site in the N-terminal domain A (site 1) that regulates neurite elongation and SV mobilization. Here, we have examined the role of phosphorylation of synapsin domain A in PTP and other forms of short-term synaptic enhancement (STE) at synapses between cultured Helix pomatia neurons. To this aim, we cloned H. pomatia synapsin (helSyn) and overexpressed GFP-tagged wild-type helSyn or site-1-mutant helSyn mutated in the presynaptic compartment of C1-B2 synapses. We found that PTP at these synapses depends both on Ca2+/calmodulin-dependent and cAMP-dependent protein kinases, and that overexpression of the non-phosphorylatable helSyn mutant, but not wild-type helSyn, specifically impairs PTP, while not altering facilitation and augmentation. Our findings show that phosphorylation of site 1 has a prominent role in the expression of PTP, thus defining a novel role for phosphorylation of synapsin domain A in short-term homosynaptic plasticity.
Figures
Similar articles
-
MAPK/Erk-dependent phosphorylation of synapsin mediates formation of functional synapses and short-term homosynaptic plasticity.J Cell Sci. 2010 Mar 15;123(Pt 6):881-93. doi: 10.1242/jcs.056846. Epub 2010 Feb 16. J Cell Sci. 2010. PMID: 20159961
-
Phosphorylation by cAMP-dependent protein kinase is essential for synapsin-induced enhancement of neurotransmitter release in invertebrate neurons.J Cell Sci. 2004 Oct 1;117(Pt 21):5145-54. doi: 10.1242/jcs.01388. J Cell Sci. 2004. PMID: 15456851
-
Intracellular injection of synapsin I induces neurotransmitter release in C1 neurons of Helix pomatia contacting a wrong target.Neuroscience. 2001;104(1):271-80. doi: 10.1016/s0306-4522(01)00063-x. Neuroscience. 2001. PMID: 11311549
-
The synapsins: key actors of synapse function and plasticity.Prog Neurobiol. 2010 Aug;91(4):313-48. doi: 10.1016/j.pneurobio.2010.04.006. Epub 2010 May 10. Prog Neurobiol. 2010. PMID: 20438797 Review.
-
The role of synapsins in neuronal development.Cell Mol Life Sci. 2010 May;67(9):1383-96. doi: 10.1007/s00018-009-0227-8. Epub 2009 Dec 25. Cell Mol Life Sci. 2010. PMID: 20035364 Free PMC article. Review.
Cited by
-
Presynaptic Short-Term Plasticity Persists in the Absence of PKC Phosphorylation of Munc18-1.J Neurosci. 2021 Sep 1;41(35):7329-7339. doi: 10.1523/JNEUROSCI.0347-21.2021. Epub 2021 Jul 21. J Neurosci. 2021. PMID: 34290081 Free PMC article.
-
Site-specific synapsin I phosphorylation participates in the expression of post-tetanic potentiation and its enhancement by BDNF.J Neurosci. 2012 Apr 25;32(17):5868-79. doi: 10.1523/JNEUROSCI.5275-11.2012. J Neurosci. 2012. PMID: 22539848 Free PMC article.
-
Molecular Mechanisms of Short-Term Plasticity: Role of Synapsin Phosphorylation in Augmentation and Potentiation of Spontaneous Glutamate Release.Front Synaptic Neurosci. 2018 Oct 30;10:33. doi: 10.3389/fnsyn.2018.00033. eCollection 2018. Front Synaptic Neurosci. 2018. PMID: 30425632 Free PMC article.
-
Selective modulation of chemical and electrical synapses of Helix neuronal networks during in vitro development.BMC Neurosci. 2013 Feb 25;14:22. doi: 10.1186/1471-2202-14-22. BMC Neurosci. 2013. PMID: 23442557 Free PMC article.
-
Short-term forms of presynaptic plasticity.Curr Opin Neurobiol. 2011 Apr;21(2):269-74. doi: 10.1016/j.conb.2011.02.003. Epub 2011 Feb 23. Curr Opin Neurobiol. 2011. PMID: 21353526 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
