

Enhanced subunit interactions with gemcitabine-5'-diphosphate inhibit ribonucleotide reductases
- PMID: 17726094
- PMCID: PMC1964847
- DOI: 10.1073/pnas.0706803104
Enhanced subunit interactions with gemcitabine-5'-diphosphate inhibit ribonucleotide reductases
Abstract
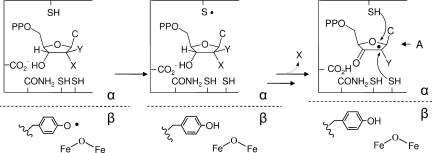
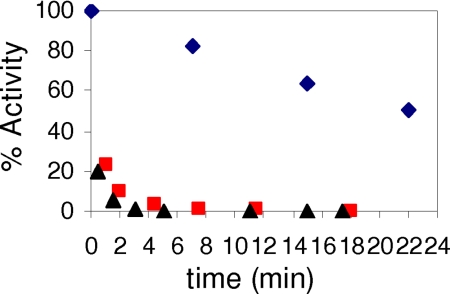
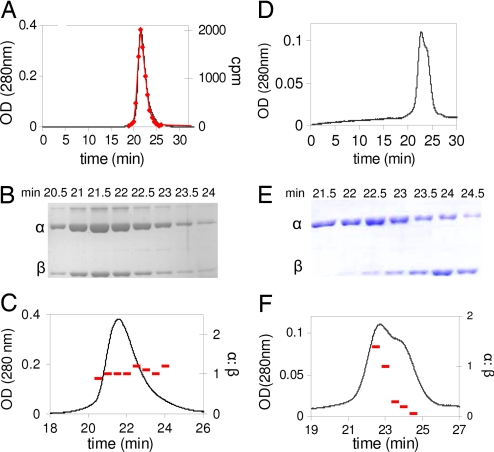
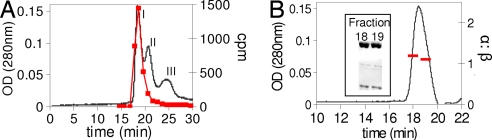
Ribonucleotide reductases (RNRs) catalyze the conversion of nucleotides to deoxynucleotides in all organisms. The class I RNRs are composed of two subunits, alpha and beta, with proposed quaternary structures of alpha2beta2, alpha6beta2, or alpha6beta6, depending on the organism. The alpha subunits bind the nucleoside diphosphate substrates and the dNTP/ATP allosteric effectors that govern specificity and turnover. The beta2 subunit houses the diferric Y* (1 radical per beta2) cofactor that is required to initiate nucleotide reduction. 2',2'-difluoro-2'-deoxycytidine (F2C) is presently used clinically in a variety of cancer treatments and the 5'-diphosphorylated F2C (F2CDP) is a potent inhibitor of RNRs. The studies with [1'-(3)H]-F2CDP and [5-(3)H]-F2CDP have established that F2CDP is a substoichiometric mechanism based inhibitor (0.5 eq F2CDP/alpha) of both the Escherichia coli and the human RNRs in the presence of reductant. Inactivation is caused by covalent labeling of RNR by the sugar of F2CDP (0.5 eq/alpha) and is accompanied by release of 0.5 eq cytosine/alpha. Inactivation also results in loss of 40% of beta2 activity. Studies using size exclusion chromatography reveal that in the E. coli RNR, an alpha2beta2 tight complex is generated subsequent to enzyme inactivation by F2CDP, whereas in the human RNR, an alpha6beta6 tight complex is generated. Isolation of these complexes establishes that the weak interactions of the subunits in the absence of nucleotides are substantially increased in the presence of F2CDP and ATP. This information and the proposed asymmetry between the interactions of alphanbetan provide an explanation for complete inactivation of RNR with substoichiometric amounts of F2CDP.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Hertel LW, Boder GB, Kroin JS, Rinzel SM, Poore GA, Todd GC, Grindey GB. Cancer Res. 1990;50:4417–4422. - PubMed
-
- Huang P, Chubb S, Hertel LW, Grindey GB, Plunkett W. Cancer Res. 1991;51:6110–6117. - PubMed
-
- Plunkett W, Huang P, Gandhi V. Nucleosides Nucleotides. 1997;16:1261–1270.
-
- Mackey JR, Mani RS, Selner M, Mowles D, Young JD, Belt JA, Crawford CR, Cass CE. Cancer Res. 1998;58:4349–4357. - PubMed
-
- Bergman AM, Pinedo HM, Peters GJ. Drug Resist Updat. 2002;5:19–33. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
