

CD200 and its receptor, CD200R, modulate bone mass via the differentiation of osteoclasts
- PMID: 17726108
- PMCID: PMC1955461
- DOI: 10.1073/pnas.0702811104
CD200 and its receptor, CD200R, modulate bone mass via the differentiation of osteoclasts
Abstract
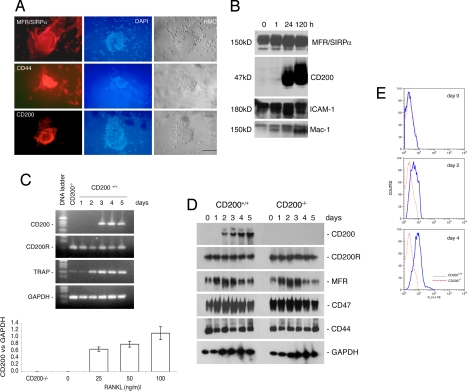
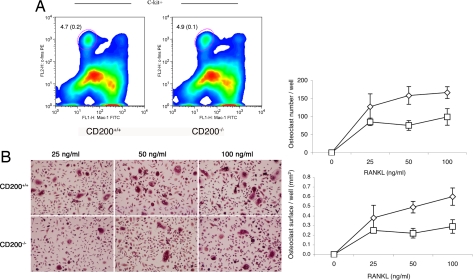
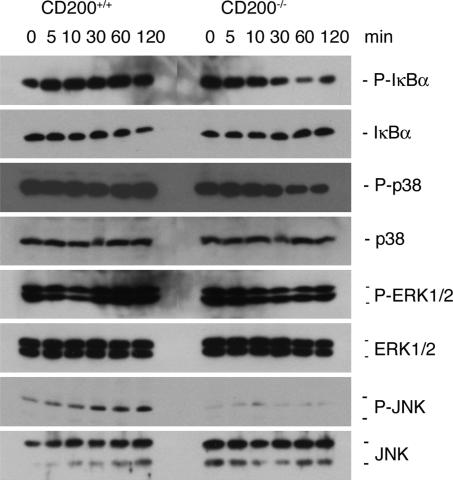
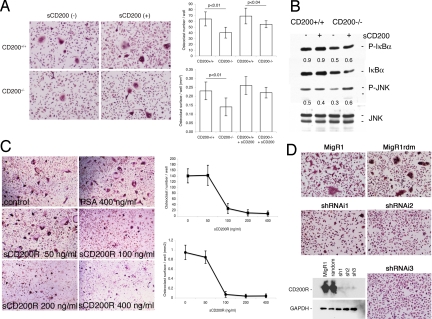
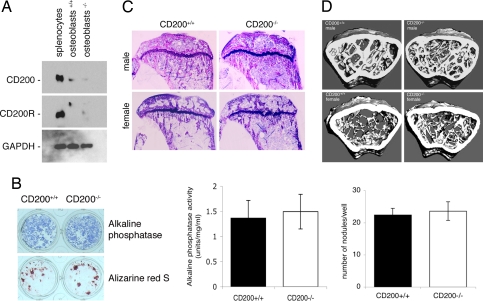
Fusion of macrophages is an essential step in the differentiation of osteoclasts, which play a central role in the development and remodeling of bone. Osteoclasts are important mediators of bone loss, which leads, for example, to osteoporosis. Macrophage fusion receptor/signal regulatory protein alpha (MFR/SIRPalpha) and its ligand CD47, which are members of the Ig superfamily (IgSF), have been implicated in the fusion of macrophages. We show that CD200, which is not expressed in cells that belong to the myeloid lineage, is strongly expressed in macrophages at the onset of fusion. By contrast, the CD200 receptor (CD200R), which, like CD200, belongs to the IgSF, is expressed only in cells that belong to the myeloid lineage, including osteoclasts, and in CD4+ T cells. Osteoclasts from CD200-/- mice differentiated at a reduced rate. Activation of the NF-kappaB and MAP kinase signaling pathways downstream of RANK, a receptor that plays a central role in the differentiation of osteoclasts, was depressed in these cells. A soluble recombinant protein that included the extracellular domain of CD200 rescued the fusion of CD200-/- macrophages and their activation downstream of RANK. Conversely, addition of a soluble recombinant protein that included the extracellular domain of CD200R or short-hairpin RNA-mediated silencing of the expression of CD200R prevented fusion. Thus CD200 engagement of the CD200R at the initiation of macrophage fusion regulated further differentiation to osteoclasts. Consistent with in vitro observations, CD200-/- mice contained fewer osteoclasts and accumulated more bone than CD200+/+ mice. The CD200-CD200R axis is therefore a putative regulator of bone mass, via the formation of osteoclasts.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
A role for the immunomodulatory molecules CD200 and CD200R in regulating bone formation.Immunol Lett. 2006 Jun 15;105(2):150-8. doi: 10.1016/j.imlet.2006.02.002. Epub 2006 Feb 28. Immunol Lett. 2006. PMID: 16530849
-
CD200R/CD200 inhibits osteoclastogenesis: new mechanism of osteoclast control by mesenchymal stem cells in human.PLoS One. 2013 Aug 5;8(8):e72831. doi: 10.1371/journal.pone.0072831. Print 2013. PLoS One. 2013. PMID: 23940819 Free PMC article.
-
Ligation of CD200R by CD200 is not required for normal murine myelopoiesis.Eur J Haematol. 2007 Nov;79(5):410-6. doi: 10.1111/j.1600-0609.2007.00920.x. Epub 2007 Sep 4. Eur J Haematol. 2007. PMID: 17803680
-
CD200/CD200R paired potent inhibitory molecules regulating immune and inflammatory responses; Part I: CD200/CD200R structure, activation, and function.Acta Medica (Hradec Kralove). 2012;55(1):12-7. doi: 10.14712/18059694.2015.68. Acta Medica (Hradec Kralove). 2012. PMID: 22696929 Review.
-
Is the CD200/CD200 receptor interaction more than just a myeloid cell inhibitory signal?Crit Rev Immunol. 2006;26(3):213-30. doi: 10.1615/critrevimmunol.v26.i3.20. Crit Rev Immunol. 2006. PMID: 16928187 Free PMC article. Review.
Cited by
-
Association of 3q13.32 variants with hip trochanter and intertrochanter bone mineral density identified by a genome-wide association study.Osteoporos Int. 2016 Nov;27(11):3343-3354. doi: 10.1007/s00198-016-3663-y. Epub 2016 Jun 16. Osteoporos Int. 2016. PMID: 27311723
-
Osteoimmunology: interactions of the bone and immune system.Endocr Rev. 2008 Jun;29(4):403-40. doi: 10.1210/er.2007-0038. Epub 2008 May 1. Endocr Rev. 2008. PMID: 18451259 Free PMC article. Review.
-
IL15RA is required for osteoblast function and bone mineralization.Bone. 2017 Oct;103:20-30. doi: 10.1016/j.bone.2017.06.003. Epub 2017 Jun 6. Bone. 2017. PMID: 28602725 Free PMC article.
-
Role of MKP-1 in osteoclasts and bone homeostasis.Am J Pathol. 2009 Oct;175(4):1564-73. doi: 10.2353/ajpath.2009.090035. Epub 2009 Sep 17. Am J Pathol. 2009. PMID: 19762714 Free PMC article.
-
STATs and macrophage fusion.JAKSTAT. 2013 Jul 1;2(3):e24777. doi: 10.4161/jkst.24777. Epub 2013 Apr 23. JAKSTAT. 2013. PMID: 24069561 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
