

Galphai generates multiple Pins activation states to link cortical polarity and spindle orientation in Drosophila neuroblasts
- PMID: 17726110
- PMCID: PMC1964812
- DOI: 10.1073/pnas.0701812104
Galphai generates multiple Pins activation states to link cortical polarity and spindle orientation in Drosophila neuroblasts
Abstract
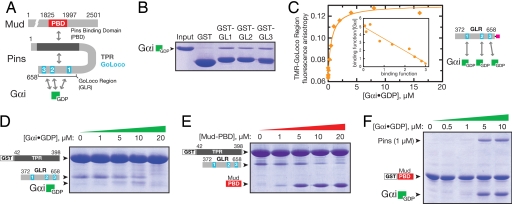
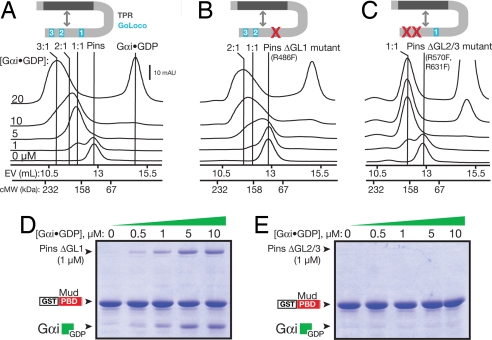
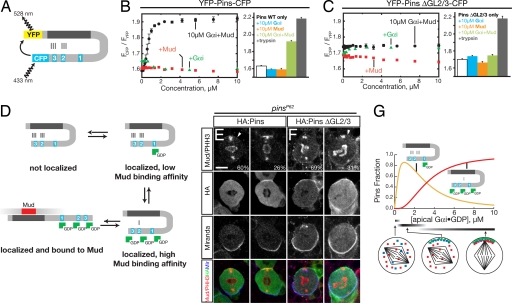
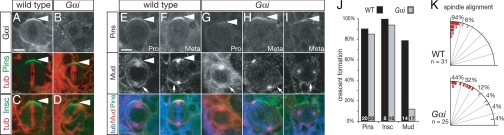
Drosophila neuroblasts divide asymmetrically by aligning their mitotic spindle with cortical cell polarity to generate distinct sibling cell types. Neuroblasts asymmetrically localize Galphai, Pins, and Mud proteins; Pins/Galphai direct cortical polarity, whereas Mud is required for spindle orientation. It is currently unknown how Galphai-Pins-Mud binding is regulated to link cortical polarity with spindle orientation. Here, we show that Pins forms a "closed" state via intramolecular GoLoco-tetratricopeptide repeat (TPR) interactions, which regulate Mud binding. Biochemical, genetic, and live imaging experiments show that Galphai binds to the first of three Pins GoLoco motifs to recruit Pins to the apical cortex without "opening" Pins or recruiting Mud. However, Galphai and Mud bind cooperatively to the Pins GoLocos 2/3 and tetratricopeptide repeat domains, respectively, thereby restricting Pins-Mud interaction to the apical cortex and fixing spindle orientation. We conclude that Pins has multiple activity states that generate cortical polarity and link it with mitotic spindle orientation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Inscuteable regulates the Pins-Mud spindle orientation pathway.PLoS One. 2012;7(1):e29611. doi: 10.1371/journal.pone.0029611. Epub 2012 Jan 10. PLoS One. 2012. PMID: 22253744 Free PMC article.
-
Canoe binds RanGTP to promote Pins(TPR)/Mud-mediated spindle orientation.J Cell Biol. 2011 Oct 31;195(3):369-76. doi: 10.1083/jcb.201102130. Epub 2011 Oct 24. J Cell Biol. 2011. PMID: 22024168 Free PMC article.
-
Robust spindle alignment in Drosophila neuroblasts by ultrasensitive activation of pins.Mol Cell. 2011 Aug 19;43(4):540-9. doi: 10.1016/j.molcel.2011.06.030. Mol Cell. 2011. PMID: 21855794 Free PMC article.
-
Epithelial polarity and spindle orientation: intersecting pathways.Philos Trans R Soc Lond B Biol Sci. 2013 Sep 23;368(1629):20130291. doi: 10.1098/rstb.2013.0291. Print 2013. Philos Trans R Soc Lond B Biol Sci. 2013. PMID: 24062590 Free PMC article. Review.
-
The cell cycle machinery and asymmetric cell division of neural progenitors in the Drosophila embryonic central nervous system.Novartis Found Symp. 2001;237:139-51; discussion 151-63. doi: 10.1002/0470846666.ch11. Novartis Found Symp. 2001. PMID: 11444041 Review.
Cited by
-
Molecular pathways regulating mitotic spindle orientation in animal cells.Development. 2013 May;140(9):1843-56. doi: 10.1242/dev.087627. Development. 2013. PMID: 23571210 Free PMC article. Review.
-
Structure of an enzyme-derived phosphoprotein recognition domain.PLoS One. 2012;7(4):e36014. doi: 10.1371/journal.pone.0036014. Epub 2012 Apr 24. PLoS One. 2012. PMID: 22545154 Free PMC article.
-
Par3 controls epithelial spindle orientation by aPKC-mediated phosphorylation of apical Pins.Curr Biol. 2010 Oct 26;20(20):1809-18. doi: 10.1016/j.cub.2010.09.032. Epub 2010 Oct 7. Curr Biol. 2010. PMID: 20933426 Free PMC article.
-
Loss of the canonical spindle orientation function in the Pins/LGN homolog AGS3.EMBO Rep. 2017 Sep;18(9):1509-1520. doi: 10.15252/embr.201643048. Epub 2017 Jul 6. EMBO Rep. 2017. PMID: 28684399 Free PMC article.
-
NuMA after 30 years: the matrix revisited.Trends Cell Biol. 2010 Apr;20(4):214-22. doi: 10.1016/j.tcb.2010.01.003. Trends Cell Biol. 2010. PMID: 20137953 Free PMC article. Review.
References
-
- Betschinger J, Knoblich JA. Curr Biol. 2004;14:R674–R685. - PubMed
-
- Cowan CR, Hyman AA. Annu Rev Cell Dev Biol. 2004;20:427–453. - PubMed
-
- Yu F, Kuo CT, Jan YN. Neuron. 2006;51:13–20. - PubMed
-
- Wodarz A. Curr Opin Cell Biol. 2005;17:475–481. - PubMed
-
- Morrison SJ, Kimble J. Nature. 2006;441:1068–1074. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
