

Insights from amphioxus into the evolution of vertebrate cartilage
- PMID: 17726517
- PMCID: PMC1950077
- DOI: 10.1371/journal.pone.0000787
Insights from amphioxus into the evolution of vertebrate cartilage
Abstract
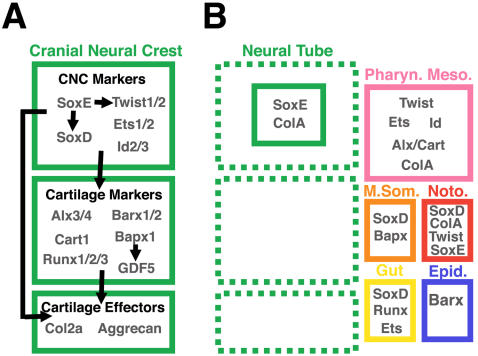
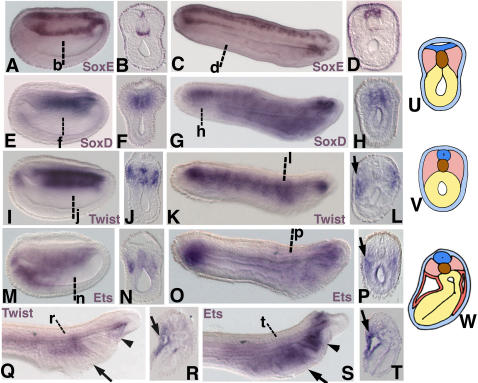
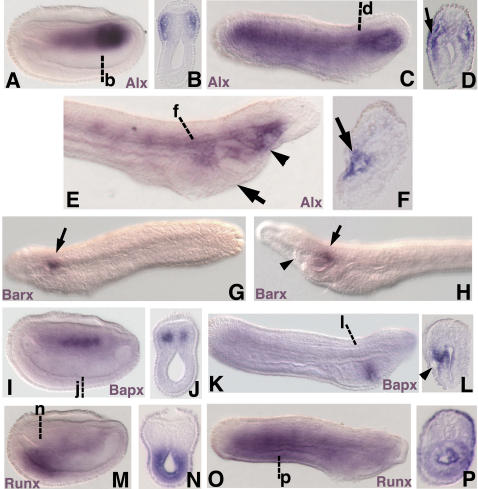
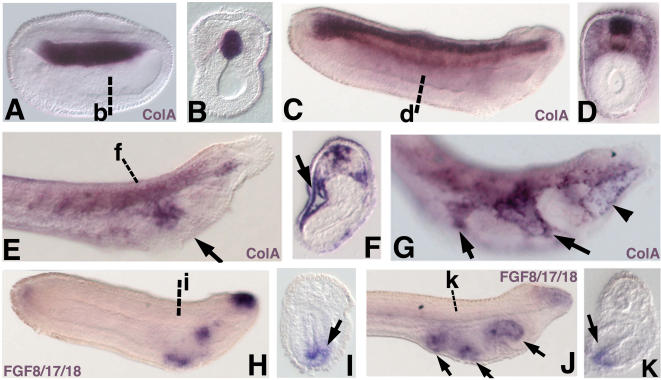
Central to the story of vertebrate evolution is the origin of the vertebrate head, a problem difficult to approach using paleontology and comparative morphology due to a lack of unambiguous intermediate forms. Embryologically, much of the vertebrate head is derived from two ectodermal tissues, the neural crest and cranial placodes. Recent work in protochordates suggests the first chordates possessed migratory neural tube cells with some features of neural crest cells. However, it is unclear how and when these cells acquired the ability to form cellular cartilage, a cell type unique to vertebrates. It has been variously proposed that the neural crest acquired chondrogenic ability by recruiting proto-chondrogenic gene programs deployed in the neural tube, pharynx, and notochord. To test these hypotheses we examined the expression of 11 amphioxus orthologs of genes involved in neural crest chondrogenesis. Consistent with cellular cartilage as a vertebrate novelty, we find that no single amphioxus tissue co-expresses all or most of these genes. However, most are variously co-expressed in mesodermal derivatives. Our results suggest that neural crest-derived cartilage evolved by serial cooption of genes which functioned primitively in mesoderm.
Conflict of interest statement
Figures
Similar articles
-
A new mechanistic scenario for the origin and evolution of vertebrate cartilage.PLoS One. 2011;6(7):e22474. doi: 10.1371/journal.pone.0022474. Epub 2011 Jul 22. PLoS One. 2011. PMID: 21799866 Free PMC article.
-
Evolution of neural crest and placodes: amphioxus as a model for the ancestral vertebrate?J Anat. 2001 Jul-Aug;199(Pt 1-2):85-98. doi: 10.1046/j.1469-7580.2001.19910085.x. J Anat. 2001. PMID: 11523831 Free PMC article. Review.
-
Insights from the amphioxus genome on the origin of vertebrate neural crest.Genome Res. 2008 Jul;18(7):1127-32. doi: 10.1101/gr.076208.108. Epub 2008 Jun 18. Genome Res. 2008. PMID: 18562679 Free PMC article.
-
An amphioxus winged helix/forkhead gene, AmphiFoxD: insights into vertebrate neural crest evolution.Dev Dyn. 2002 Nov;225(3):289-97. doi: 10.1002/dvdy.10173. Dev Dyn. 2002. PMID: 12412011
-
Origins and plasticity of neural crest cells and their roles in jaw and craniofacial evolution.Int J Dev Biol. 2003;47(7-8):541-53. Int J Dev Biol. 2003. PMID: 14756330 Review.
Cited by
-
The phylum Vertebrata: a case for zoological recognition.Zoological Lett. 2018 Dec 26;4:32. doi: 10.1186/s40851-018-0114-y. eCollection 2018. Zoological Lett. 2018. PMID: 30607258 Free PMC article. Review.
-
Patterning of cartilaginous condensations in the developing facial skeleton.Dev Biol. 2022 Jun;486:44-55. doi: 10.1016/j.ydbio.2022.03.010. Epub 2022 Mar 28. Dev Biol. 2022. PMID: 35358504 Free PMC article.
-
A new mechanistic scenario for the origin and evolution of vertebrate cartilage.PLoS One. 2011;6(7):e22474. doi: 10.1371/journal.pone.0022474. Epub 2011 Jul 22. PLoS One. 2011. PMID: 21799866 Free PMC article.
-
Gene profiling of head mesoderm in early zebrafish development: insights into the evolution of cranial mesoderm.Evodevo. 2019 Jul 6;10:14. doi: 10.1186/s13227-019-0128-3. eCollection 2019. Evodevo. 2019. PMID: 31312422 Free PMC article.
-
Linking Vertebrate Gene Duplications to the New Head Hypothesis.Biology (Basel). 2023 Sep 6;12(9):1213. doi: 10.3390/biology12091213. Biology (Basel). 2023. PMID: 37759612 Free PMC article. Review.
References
-
- Couly GF, Coltey PM, Le Douarin NM. The triple origin of skull in higher vertebrates: a study in quail-chick chimeras. Development. 1993;117:409–429. - PubMed
-
- Meulemans D, Bronner-Fraser M. Central role of gene cooption in neural crest evolution. J Exp Zoolog B Mol Dev Evol. 2005;304:298–303. - PubMed
-
- Jeffery WR, Strickler AG, Yamamoto Y. Migratory neural crest-like cells form body pigmentation in a urochordate embryo. Nature. 2004;431:696–699. - PubMed
-
- Gans C, Northcutt RG. Neural Crest and the Origin of Vertebrates-a New Head. Science. 1983;220:268–274. - PubMed
-
- Northcutt RG, Gans C. The genesis of neural crest and epidermal placodes: a reinterpretation of vertebrate origins. Q Rev Biol. 1983;58:1–28. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
