

Inhibition of primordial germ cell proliferation by the medaka male determining gene Dmrt I bY
- PMID: 17760954
- PMCID: PMC2034567
- DOI: 10.1186/1471-213X-7-99
Inhibition of primordial germ cell proliferation by the medaka male determining gene Dmrt I bY
Abstract
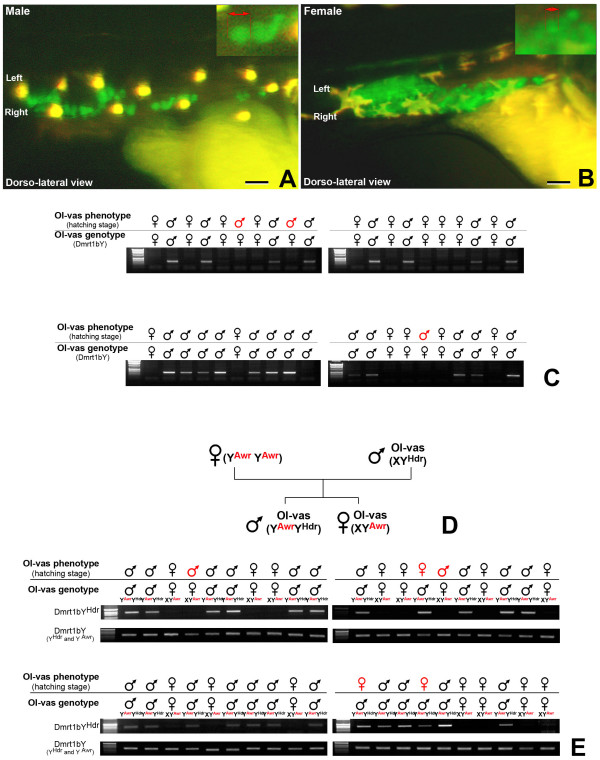
Background: Dmrt I is a highly conserved gene involved in the determination and early differentiation phase of the primordial gonad in vertebrates. In the fish medaka dmrt I bY, a functional duplicate of the autosomal dmrt I a gene on the Y-chromosome, has been shown to be the master regulator of male gonadal development, comparable to Sry in mammals. In males mRNA and protein expression was observed before morphological sex differentiation in the somatic cells surrounding primordial germ cells (PGCs) of the gonadal anlage and later on exclusively in Sertoli cells. This suggested a role for dmrt I bY during male gonad and germ cell development.
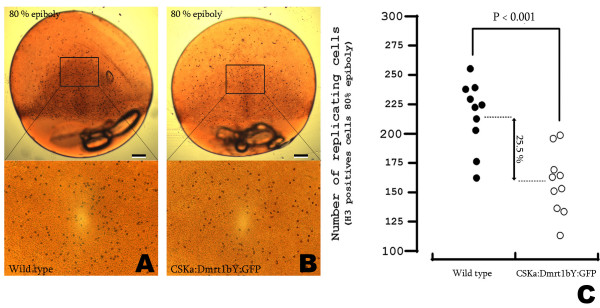
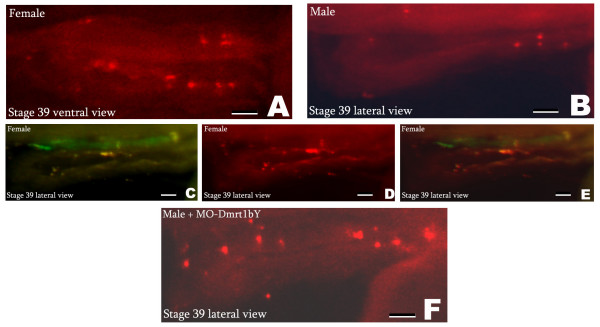
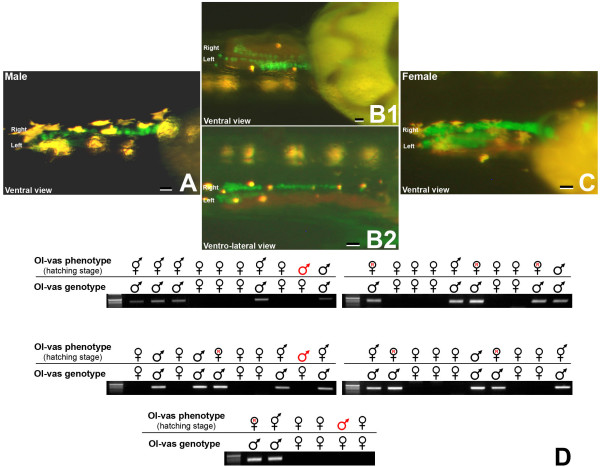
Results: We provide functional evidence that expression of dmrt I bY leads to negative regulation of PGC proliferation. Flow cytometric measurements revealed a G2 arrest of dmrt I bY expressing cells. Interestingly, also non-transfected cells displayed a significantly lower fraction of proliferating cells, pointing to a possible non-cell autonomous action of dmrt I bY. Injection of antisense morpholinos led to an increase of PGCs in genetically male embryos due to loss of proliferation inhibition.
Conclusion: In medaka, dmrt I bY mediates a mitotic arrest of PGCs in males prior to testes differentiation at the sex determination stage. This occurs possibly via a cross-talk of Sertoli cells and PGCs.
Figures



Similar articles
-
Two DM domain genes, DMY and DMRT1, involved in testicular differentiation and development in the medaka, Oryzias latipes.Dev Dyn. 2004 Nov;231(3):518-26. doi: 10.1002/dvdy.20158. Dev Dyn. 2004. PMID: 15376325
-
Cross talk between germ cells and gonadal somatic cells is critical for sex differentiation of the gonads in the teleost fish, medaka (Oryzias latipes).Dev Growth Differ. 2008 May;50(4):273-8. doi: 10.1111/j.1440-169X.2008.01015.x. Dev Growth Differ. 2008. PMID: 18366386 Review.
-
Germ cells are not the primary factor for sexual fate determination in goldfish.Dev Biol. 2012 Oct 1;370(1):98-109. doi: 10.1016/j.ydbio.2012.07.010. Epub 2012 Jul 21. Dev Biol. 2012. PMID: 22824426
-
Regulatory back-up circuit of medaka Wt1 co-orthologs ensures PGC maintenance.Dev Biol. 2009 Jan 1;325(1):179-88. doi: 10.1016/j.ydbio.2008.10.009. Epub 2008 Nov 1. Dev Biol. 2009. PMID: 18992736
-
Guidance of primordial germ cell migration.Curr Opin Cell Biol. 2004 Apr;16(2):169-73. doi: 10.1016/j.ceb.2004.01.004. Curr Opin Cell Biol. 2004. PMID: 15196560 Review.
Cited by
-
Dmy initiates masculinity by altering Gsdf/Sox9a2/Rspo1 expression in medaka (Oryzias latipes).Sci Rep. 2016 Jan 25;6:19480. doi: 10.1038/srep19480. Sci Rep. 2016. PMID: 26806354 Free PMC article.
-
In-depth investigation of genome to refine QTL positions for spontaneous sex-reversal in XX rainbow trout.PLoS One. 2025 May 7;20(5):e0313464. doi: 10.1371/journal.pone.0313464. eCollection 2025. PLoS One. 2025. PMID: 40333945 Free PMC article.
-
dmrt1 Is Responsible for Androgen-Induced Masculinization in Nile Tilapia.Genes (Basel). 2024 Sep 23;15(9):1238. doi: 10.3390/genes15091238. Genes (Basel). 2024. PMID: 39336829 Free PMC article.
-
Dmrt genes in the development and evolution of sexual dimorphism.Trends Genet. 2012 Apr;28(4):175-84. doi: 10.1016/j.tig.2012.02.002. Epub 2012 Mar 14. Trends Genet. 2012. PMID: 22425532 Free PMC article. Review.
-
Germline stem cells in human.Signal Transduct Target Ther. 2022 Oct 2;7(1):345. doi: 10.1038/s41392-022-01197-3. Signal Transduct Target Ther. 2022. PMID: 36184610 Free PMC article. Review.
References
-
- Raymond CS, Parker ED, Kettlewell JR, Brown LG, Page DC, Kusz K, Jaruzelska J, Reinberg Y, Flejter WL, Bardwell VJ, Hirsch B, Zarkower D. A region of human chromosome 9p required for testis development contains two genes related to known sexual regulators. Hum Mol Genet. 1999;8:989–96. doi: 10.1093/hmg/8.6.989. - DOI - PubMed
-
- Nanda I, Kondo M, Hornung U, Asakawa S, Winkler C, Shimizu A, Shan Z, Haaf T, Shimizu N, Shima A, Schmid M, Schartl M. A duplicated copy of DMRT1 in the sex-determining region of the Y chromosome of the medaka, Oryzias latipes. Proc Natl Acad Sci USA. 2002;99:11778–83. doi: 10.1073/pnas.182314699. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
