

Ethanol preconditioning protects against ischemia/reperfusion-induced brain damage: role of NADPH oxidase-derived ROS
- PMID: 17761301
- PMCID: PMC2173699
- DOI: 10.1016/j.freeradbiomed.2007.06.018
Ethanol preconditioning protects against ischemia/reperfusion-induced brain damage: role of NADPH oxidase-derived ROS
Abstract
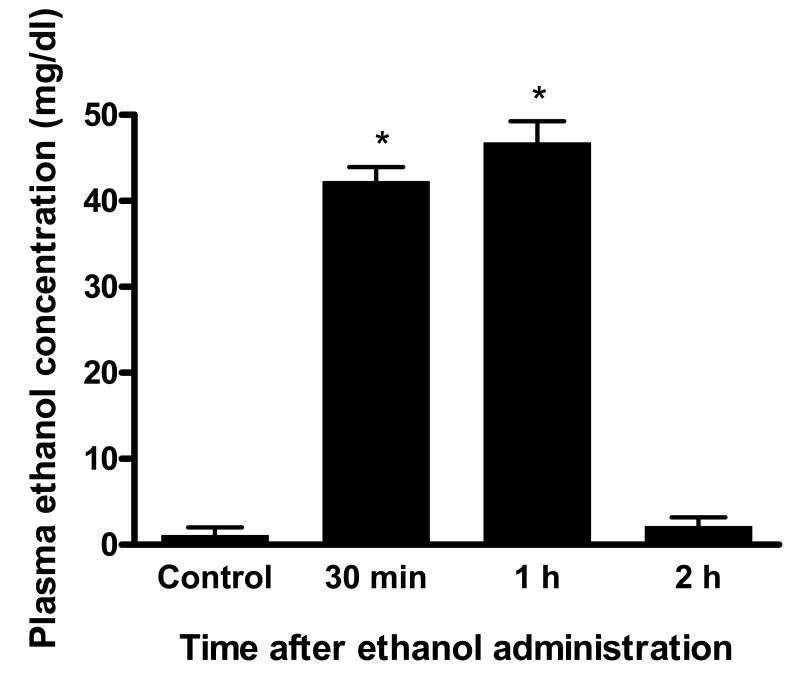
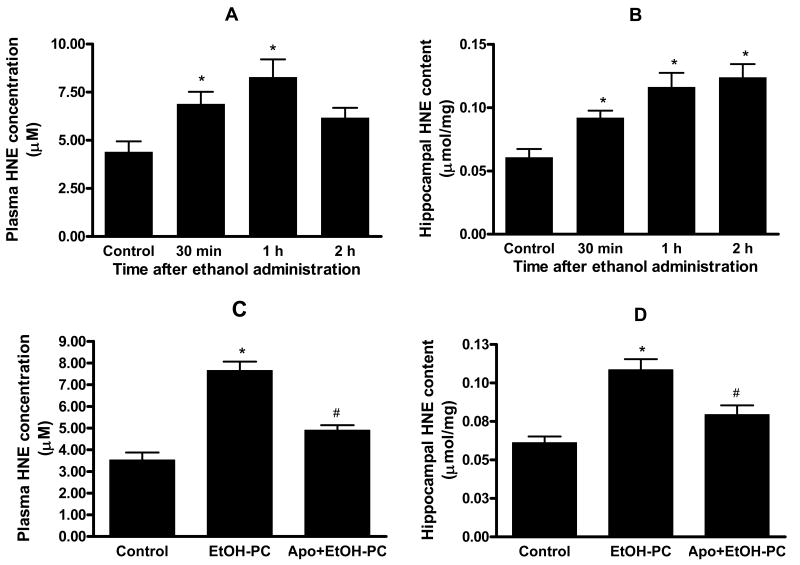
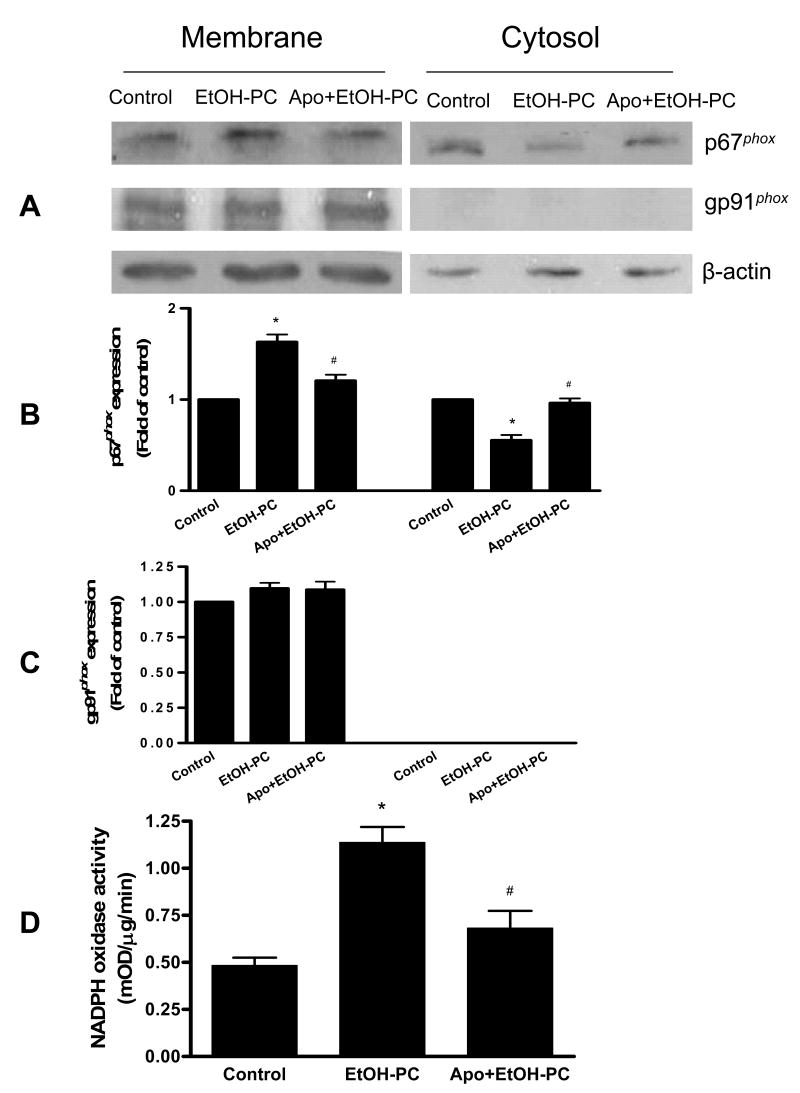
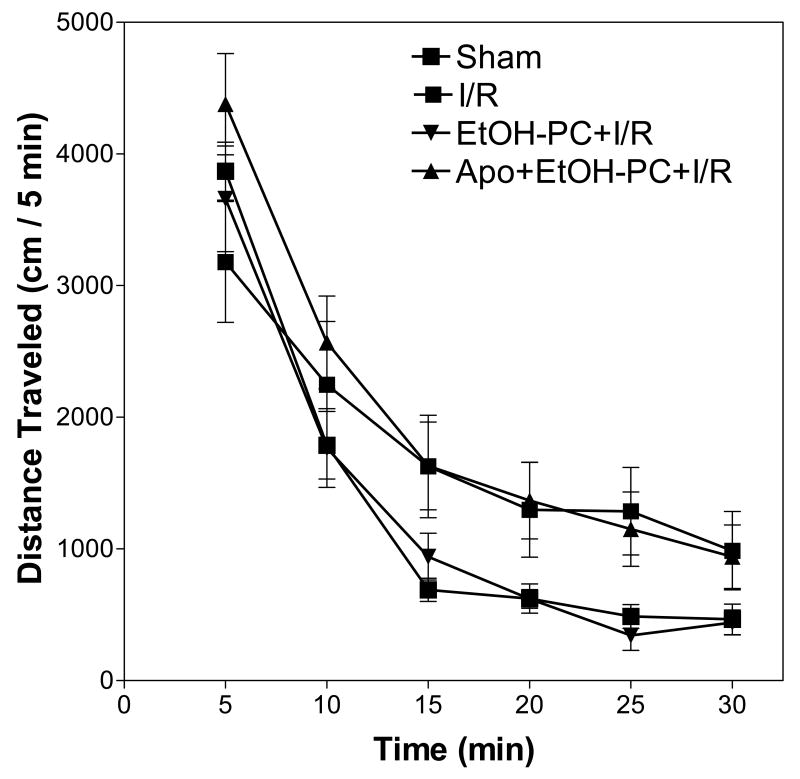
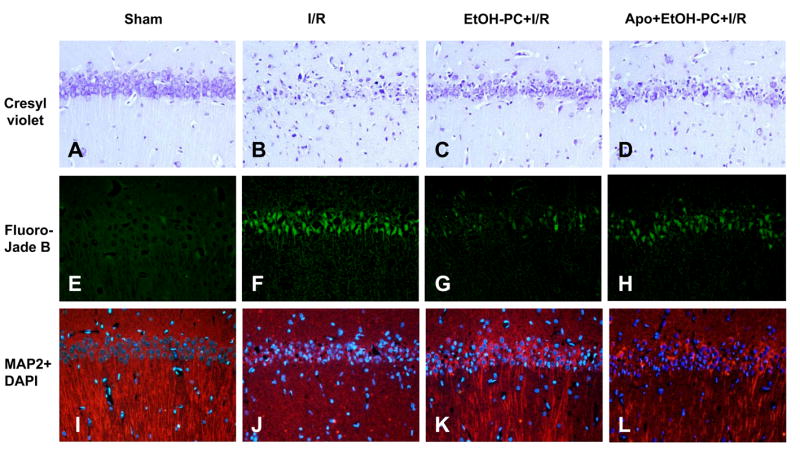
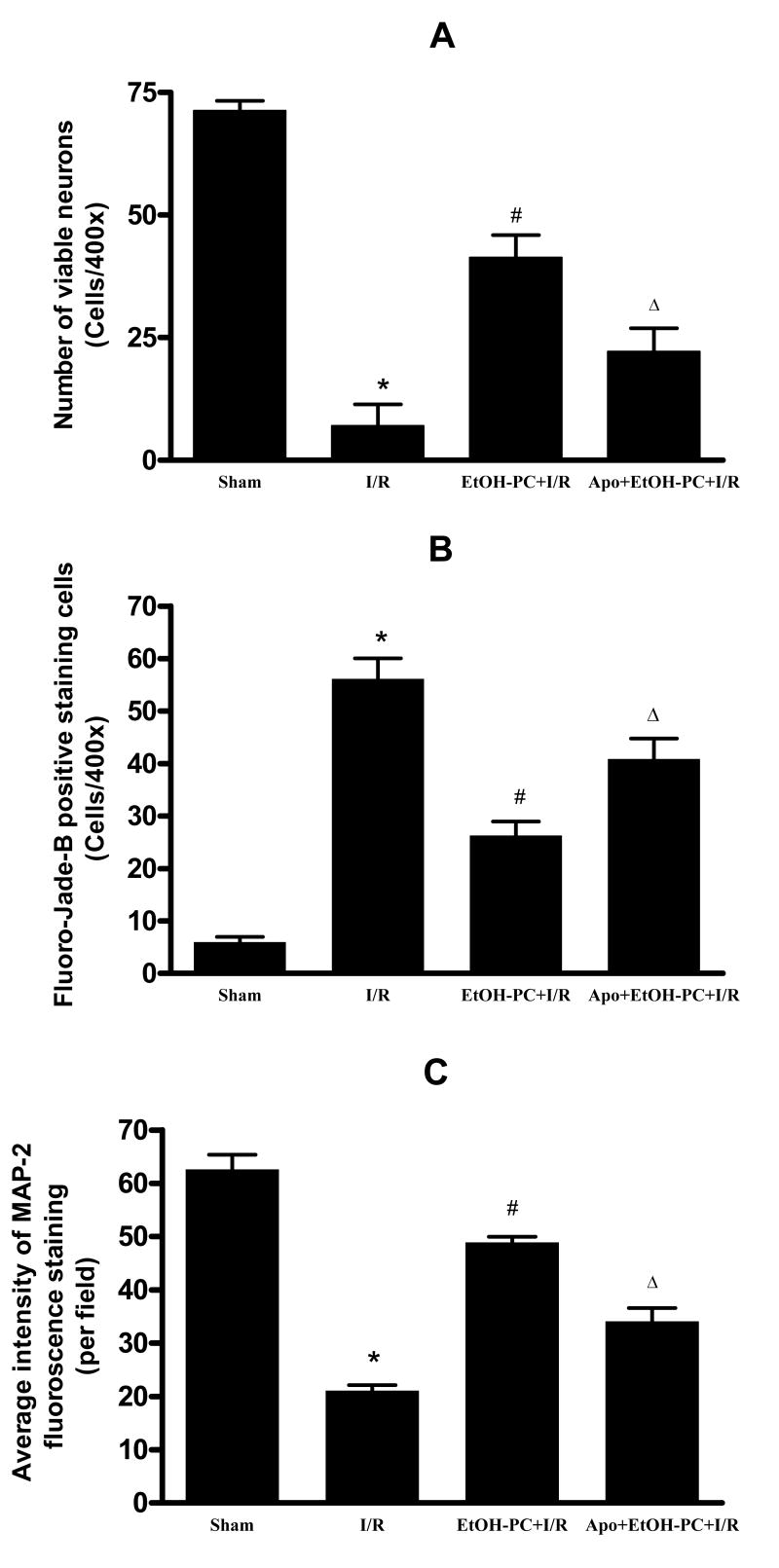
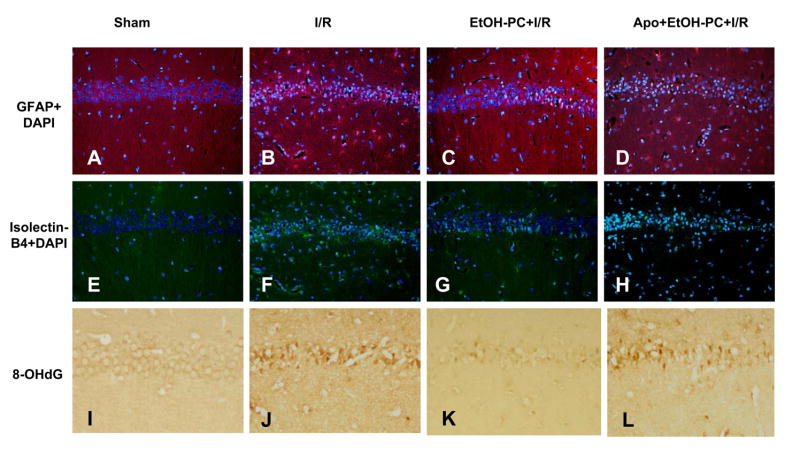
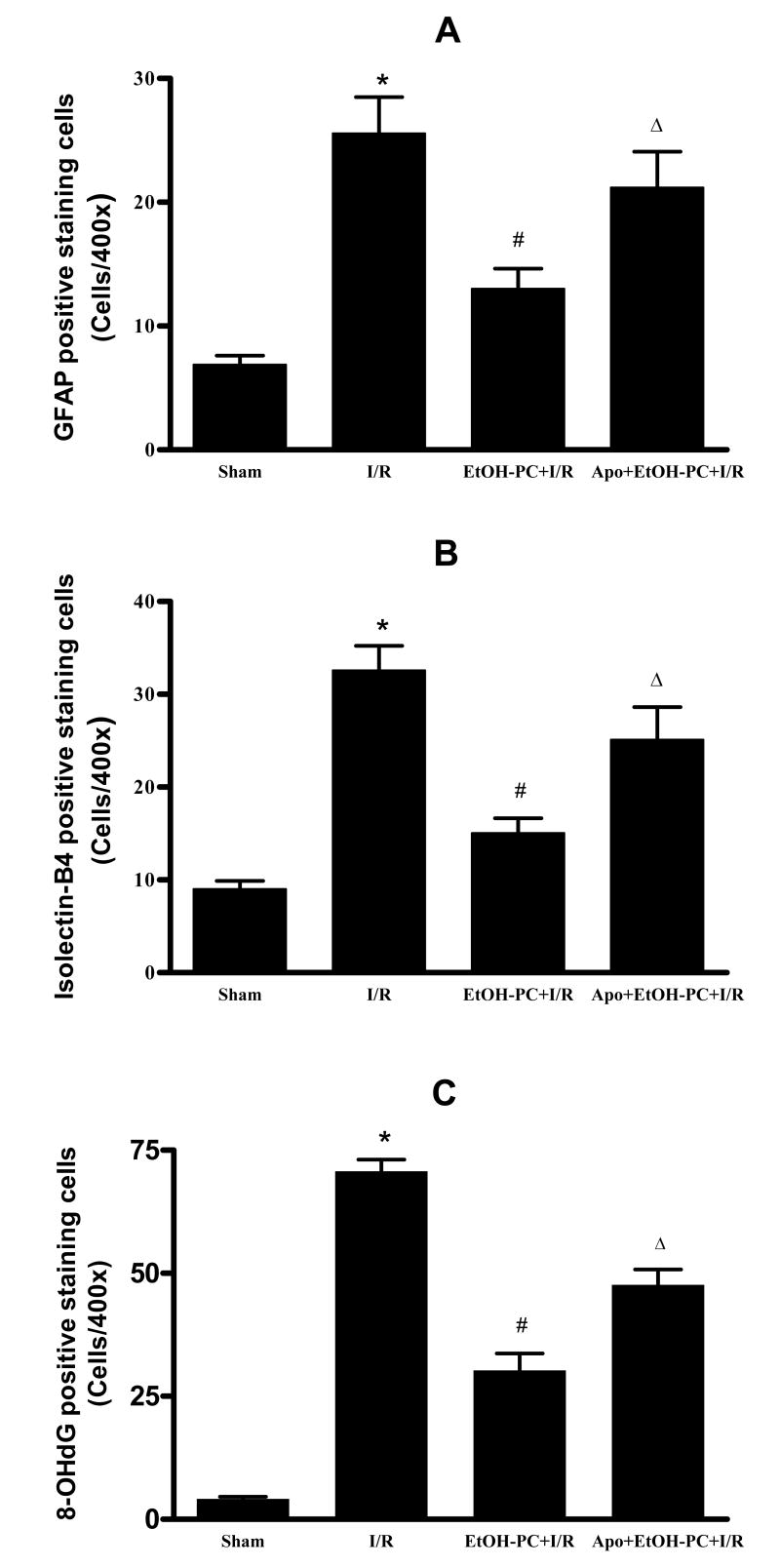
Ethanol preconditioning (EtOH-PC) refers to a phenomenon in which tissues are protected from the deleterious effects of ischemia/reperfusion (I/R) by prior ingestion of ethanol at low to moderate levels. In this study, we tested whether prior (24 h) administration of ethanol as a single bolus that produced a peak plasma concentration of 42-46 mg/dl in gerbils would offer protective effects against neuronal damage due to cerebral I/R. In addition, we also tested whether reactive oxygen species (ROS) derived from NADPH oxidase played a role as initiators of these putative protective effects. Groups of gerbils were administered either ethanol or the same volume of water by gavage 24 h before transient global cerebral ischemia induced by occlusion of both common carotid arteries for 5 min. In some experiments, apocynin, a specific inhibitor of NADPH oxidase, was administered (5 mg/kg body wt, i.p.) 10 min before ethanol administration. EtOH-PC ameliorated behavioral deficit induced by cerebral I/R and protected the brain against I/R-induced delayed neuronal death, neuronal and dendritic degeneration, oxidative DNA damage, and glial cell activation. These beneficial effects were attenuated by apocynin treatment coincident with ethanol administration. Ethanol ingestion was associated with translocation of the NADPH oxidase subunit p67(phox) from hippocampal cytosol fraction to membrane, increased NADPH oxidase activity in hippocampus within the first hour after gavage, and increased lipid peroxidation (4-hydroxy-2-nonenal) in plasma and hippocampus within the first 2 h after gavage. These effects were also inhibited by concomitant apocynin treatment. Our data are consistent with the hypothesis that antecedent ethanol ingestion at socially relevant levels induces neuroprotective effects in I/R by a mechanism that is triggered by ROS produced through NADPH oxidase. Our results further suggest the possibility that preconditioning with other pharmacological agents that induce a mild oxidative stress may have similar therapeutic value for suppressing stroke-mediated damage in brain.
Figures
References
-
- Truelsen T, Gronbaek M. Wine consumption and cerebrovascular disease mortality in Spain. Stroke. 1999;30:186–188. - PubMed
-
- German JB, Walzem RL. The health benefits of wine. Annu Rev Nutr. 2000;20:561–593. - PubMed
-
- Rakotovao A, Berthonneche C, Guiraud A, de Lorgeril M, Salen P, de Leiris J, Boucher F. Ethanol, wine, and experimental cardioprotection in ischemia/reperfusion: role of the prooxidant/antioxidant balance. Antioxid Redox Signal. 2004;6:431–438. - PubMed
-
- Ray PS, Maulik G, Cordis GA, Bertelli AA, Bertelli A, Das DK. The red wine antioxidant resveratrol protects isolated rat hearts from ischemia reperfusion injury. Free Radic Biol Med. 1999;27:160–169. - PubMed
-
- Wang Q, Xu J, Rottinghaus GE, Simonyi A, Lubahn D, Sun GY, Sun AY. Resveratrol protects against global cerebral ischemic injury in gerbils. Brain Res. 2002;958:439–447. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
