

AtPRD1 is required for meiotic double strand break formation in Arabidopsis thaliana
- PMID: 17762870
- PMCID: PMC2230667
- DOI: 10.1038/sj.emboj.7601815
AtPRD1 is required for meiotic double strand break formation in Arabidopsis thaliana
Abstract
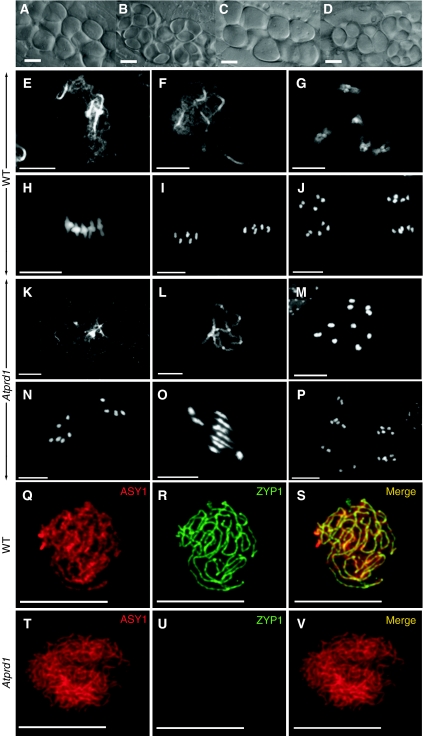
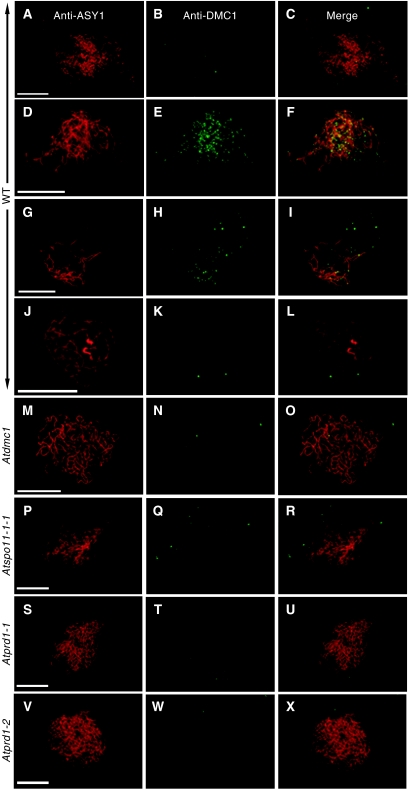
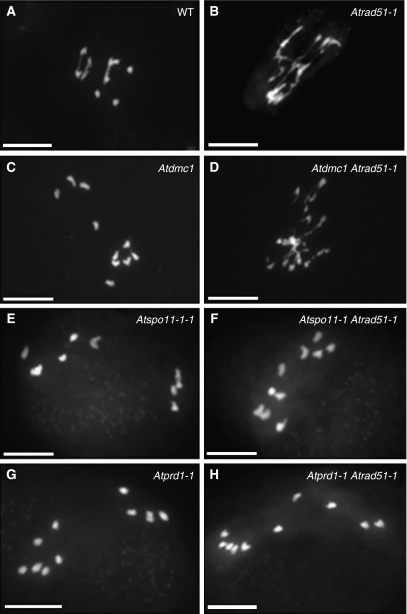
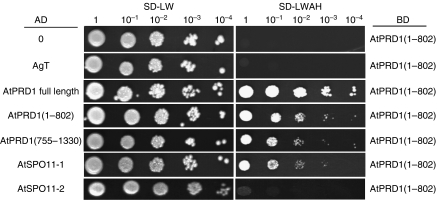
The initiation of meiotic recombination by the formation of DNA double-strand breaks (DSBs) catalysed by the Spo11 protein is strongly evolutionary conserved. In Saccharomyces cerevisiae, Spo11 requires nine other proteins for meiotic DSB formation, but, unlike Spo11, few of these proteins seem to be conserved across kingdoms. In order to investigate this recombination step in higher eukaryotes, we have isolated a new gene, AtPRD1, whose mutation affects meiosis in Arabidopsis thaliana. In Atprd1 mutants, meiotic recombination rates fall dramatically, early recombination markers (e.g., DMC1 foci) are absent, but meiosis progresses until achiasmatic univalents are formed. Besides, Atprd1 mutants suppress DSB repair defects of a large range of meiotic mutants, showing that AtPRD1 is involved in meiotic recombination and is required for meiotic DSB formation. Furthermore, we showed that AtPRD1 and AtSPO11-1 interact in a yeast two-hybrid assay, suggesting that AtPRD1 could be a partner of AtSPO11-1. Moreover, our study reveals similarity between AtPRD1 and the mammalian protein Mei1, suggesting that AtPRD1 could be a Mei1 functional homologue.
Figures

Similar articles
-
A high throughput genetic screen identifies new early meiotic recombination functions in Arabidopsis thaliana.PLoS Genet. 2009 Sep;5(9):e1000654. doi: 10.1371/journal.pgen.1000654. Epub 2009 Sep 18. PLoS Genet. 2009. PMID: 19763177 Free PMC article.
-
The Arabidopsis thaliana DSB formation (AtDFO) gene is required for meiotic double-strand break formation.Plant J. 2012 Oct;72(2):271-81. doi: 10.1111/j.1365-313X.2012.05075.x. Epub 2012 Jul 26. Plant J. 2012. PMID: 22694475
-
MTOPVIB interacts with AtPRD1 and plays important roles in formation of meiotic DNA double-strand breaks in Arabidopsis.Sci Rep. 2017 Aug 30;7(1):10007. doi: 10.1038/s41598-017-10270-9. Sci Rep. 2017. PMID: 28855712 Free PMC article.
-
DNA double-strand breaks in meiosis: checking their formation, processing and repair.DNA Repair (Amst). 2009 Sep 2;8(9):1127-38. doi: 10.1016/j.dnarep.2009.04.005. Epub 2009 May 22. DNA Repair (Amst). 2009. PMID: 19464965 Review.
-
Modulating and targeting meiotic double-strand breaks in Saccharomyces cerevisiae.Methods Mol Biol. 2009;557:27-33. doi: 10.1007/978-1-59745-527-5_3. Methods Mol Biol. 2009. PMID: 19799174 Review.
Cited by
-
Crossover interference mechanism: New lessons from plants.Front Cell Dev Biol. 2023 May 19;11:1156766. doi: 10.3389/fcell.2023.1156766. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 37274744 Free PMC article. Review.
-
Conservation and divergence of meiotic DNA double strand break forming mechanisms in Arabidopsis thaliana.Nucleic Acids Res. 2021 Sep 27;49(17):9821-9835. doi: 10.1093/nar/gkab715. Nucleic Acids Res. 2021. PMID: 34458909 Free PMC article.
-
Identification of QTNs Associated With Flowering Time, Maturity, and Plant Height Traits in Linum usitatissimum L. Using Genome-Wide Association Study.Front Genet. 2022 Jun 14;13:811924. doi: 10.3389/fgene.2022.811924. eCollection 2022. Front Genet. 2022. PMID: 35774513 Free PMC article.
-
The plant early recombinosome: a high security complex to break DNA during meiosis.Plant Reprod. 2024 Dec;37(4):421-440. doi: 10.1007/s00497-024-00509-7. Epub 2024 Sep 27. Plant Reprod. 2024. PMID: 39331138 Free PMC article. Review.
-
Targeted induction of meiotic double-strand breaks reveals chromosomal domain-dependent regulation of Spo11 and interactions among potential sites of meiotic recombination.Nucleic Acids Res. 2008 Feb;36(3):984-97. doi: 10.1093/nar/gkm1082. Epub 2007 Dec 20. Nucleic Acids Res. 2008. PMID: 18096626 Free PMC article.
References
-
- Akutsu N, Iijima K, Hinata T, Tauchi H (2007) Characterization of the plant homolog of Nijmegen breakage syndrome 1: involvement in DNA repair and recombination. Biochem Biophys Res Commun 353: 394–398 - PubMed
-
- Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P, Stevenson DK, Zimmerman J, Barajas P, Cheuk R, Gadrinab C, Heller C, Jeske A, Koesema E, Meyers CC, Parker H, Prednis L, Ansari Y, Choy N, Deen H, Geralt M, Hazari N, Hom E, Karnes M, Mulholland C, Ndubaku R, Schmidt I, Guzman P, Aguilar-Henonin L, Schmid M, Weigel D, Carter DE, Marchand T, Risseeuw E, Brogden D, Zeko A, Crosby WL, Berry CC, Ecker JR (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657 - PubMed
-
- Armstrong SJ, Caryl AP, Jones GH, Franklin FC (2002) Asy1, a protein required for meiotic chromosome synapsis, localizes to axis-associated chromatin in Arabidopsis and Brassica. J Cell Sci 115: 3645–3655 - PubMed
-
- Arora C, Kee K, Maleki S, Keeney S (2004) Antiviral protein Ski8 is a direct partner of Spo11 in meiotic DNA break formation, independent of its cytoplasmic role in RNA metabolism. Mol Cell 13: 549–559 - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
