

N-Acylethanolamine metabolism interacts with abscisic acid signaling in Arabidopsis thaliana seedlings
- PMID: 17766402
- PMCID: PMC2002614
- DOI: 10.1105/tpc.106.048702
N-Acylethanolamine metabolism interacts with abscisic acid signaling in Arabidopsis thaliana seedlings
Abstract
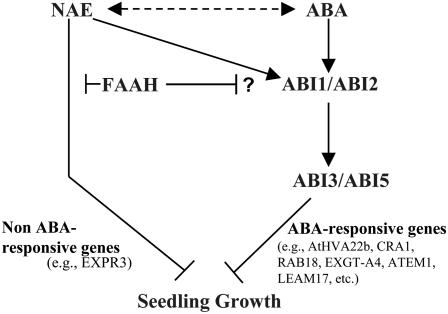
N-Acylethanolamines (NAEs) are bioactive acylamides that are present in a wide range of organisms. In plants, NAEs are generally elevated in desiccated seeds, suggesting that they may play a role in seed physiology. NAE and abscisic acid (ABA) levels were depleted during seed germination, and both metabolites inhibited the growth of Arabidopsis thaliana seedlings within a similar developmental window. Combined application of low levels of ABA and NAE produced a more dramatic reduction in germination and growth than either compound alone. Transcript profiling and gene expression studies in NAE-treated seedlings revealed elevated transcripts for a number of ABA-responsive genes and genes typically enriched in desiccated seeds. The levels of ABI3 transcripts were inversely associated with NAE-modulated growth. Overexpression of the Arabidopsis NAE degrading enzyme fatty acid amide hydrolase resulted in seedlings that were hypersensitive to ABA, whereas the ABA-insensitive mutants, abi1-1, abi2-1, and abi3-1, exhibited reduced sensitivity to NAE. Collectively, our data indicate that an intact ABA signaling pathway is required for NAE action and that NAE may intersect the ABA pathway downstream from ABA. We propose that NAE metabolism interacts with ABA in the negative regulation of seedling development and that normal seedling establishment depends on the reduction of the endogenous levels of both metabolites.
Figures








Similar articles
-
N-acylethanolamine (NAE) inhibits growth in Arabidopsis thaliana seedlings via ABI3-dependent and -independent pathways.Plant Signal Behav. 2011 May;6(5):671-9. doi: 10.4161/psb.6.5.14704. Epub 2011 May 1. Plant Signal Behav. 2011. PMID: 21633189 Free PMC article.
-
Lipoxygenase-derived 9-hydro(pero)xides of linoleoylethanolamide interact with ABA signaling to arrest root development during Arabidopsis seedling establishment.Plant J. 2015 Apr;82(2):315-27. doi: 10.1111/tpj.12821. Plant J. 2015. PMID: 25752187
-
Arabidopsis WRKY6 Transcription Factor Acts as a Positive Regulator of Abscisic Acid Signaling during Seed Germination and Early Seedling Development.PLoS Genet. 2016 Feb 1;12(2):e1005833. doi: 10.1371/journal.pgen.1005833. eCollection 2016 Feb. PLoS Genet. 2016. PMID: 26829043 Free PMC article.
-
Light and abscisic acid interplay in early seedling development.New Phytol. 2021 Jan;229(2):763-769. doi: 10.1111/nph.16963. Epub 2020 Oct 25. New Phytol. 2021. PMID: 32984965 Review.
-
N-Acylethanolamines and related compounds: aspects of metabolism and functions.Plant Sci. 2012 Mar;184:129-40. doi: 10.1016/j.plantsci.2011.12.015. Epub 2011 Dec 24. Plant Sci. 2012. PMID: 22284717 Review.
Cited by
-
Fatty acid amide hydrolase and 9-lipoxygenase modulate cotton seedling growth by ethanolamide oxylipin levels.Plant Physiol. 2023 Feb 12;191(2):1234-1253. doi: 10.1093/plphys/kiac556. Plant Physiol. 2023. PMID: 36472510 Free PMC article.
-
N-Acylated phospholipid metabolism and seedling growth: insights from lipidomics studies in Arabidopsis.Plant Signal Behav. 2012 Sep 1;7(9):1200-2. doi: 10.4161/psb.21314. Epub 2012 Aug 17. Plant Signal Behav. 2012. PMID: 22899060 Free PMC article.
-
Disruption of the Arabidopsis CGI-58 homologue produces Chanarin-Dorfman-like lipid droplet accumulation in plants.Proc Natl Acad Sci U S A. 2010 Oct 12;107(41):17833-8. doi: 10.1073/pnas.0911359107. Epub 2010 Sep 27. Proc Natl Acad Sci U S A. 2010. PMID: 20876112 Free PMC article.
-
Population genomics demystifies the defoliation phenotype in the plant pathogen Verticillium dahliae.New Phytol. 2019 Apr;222(2):1012-1029. doi: 10.1111/nph.15672. Epub 2019 Feb 25. New Phytol. 2019. PMID: 30609067 Free PMC article.
-
Antibacterial Activity and Mechanism of Three Root Exudates from Mulberry Seedlings against Ralstonia pseudosolanacearum.Plants (Basel). 2024 Feb 8;13(4):482. doi: 10.3390/plants13040482. Plants (Basel). 2024. PMID: 38498445 Free PMC article.
References
-
- Assmann, S.M. (2005). G protein signaling in the regulation of Arabidopsis seed germination. Sci. STKE 2005 cm11. - PubMed
-
- Bassel, G.W., Mullen, R.T., and Bewley, J.D. (2006). ABI3 expression ceases following, but not during, germination of tomato and Arabidopsis seeds. J. Exp. Bot. 57 1291–1297. - PubMed
-
- Blancaflor, E.B., and Chapman, K.D. (2006). Similarities between endocannabinoid signaling in animal systems and N-acylethanolamine metabolism in plants. In Communication in Plants, F. Baluska, S. Mancuso, and D. Volkmann, eds (Berlin: Springer-Verlag), pp. 205–219.
-
- Blancaflor, E.B., Hou, G., and Chapman, K.D. (2003). Elevated levels of N-lauroylethanolamine, an endogenous constituent of desiccated seeds, disrupt normal root development in Arabidopsis thaliana seedlings. Planta 217 206–217. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
