

Impact of pH on lactate formation and utilization by human fecal microbial communities
- PMID: 17766450
- PMCID: PMC2075063
- DOI: 10.1128/AEM.00508-07
Impact of pH on lactate formation and utilization by human fecal microbial communities
Abstract
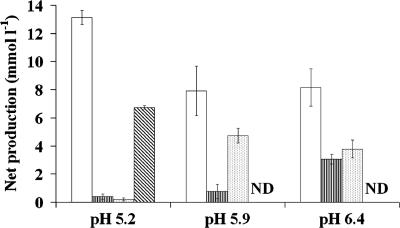
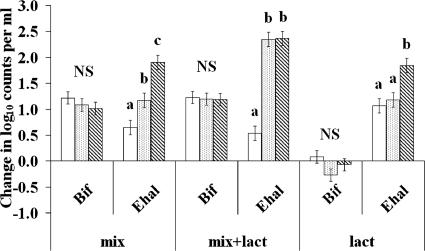
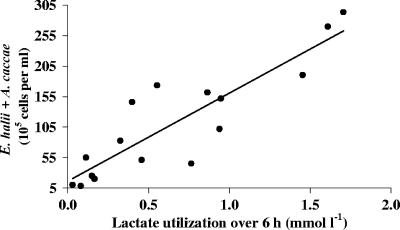
The human intestine harbors both lactate-producing and lactate-utilizing bacteria. Lactate is normally present at <3 mmol liter(-1) in stool samples from healthy adults, but concentrations up to 100 mmol liter(-1) have been reported in gut disorders such as ulcerative colitis. The effect of different initial pH values (5.2, 5.9, and 6.4) upon lactate metabolism was studied with fecal inocula from healthy volunteers, in incubations performed with the addition of dl-lactate, a mixture of polysaccharides (mainly starch), or both. Propionate and butyrate formation occurred at pH 6.4; both were curtailed at pH 5.2, while propionate but not butyrate formation was inhibited at pH 5.9. With the polysaccharide mix, lactate accumulation occurred only at pH 5.2, but lactate production, estimated using l-[U-(13)C]lactate, occurred at all three pH values. Lactate was completely utilized within 24 h at pH 5.9 and 6.4 but not at pH 5.2. At pH 5.9, more butyrate than propionate was formed from l-[U-(13)C]lactate in the presence of polysaccharides, but propionate, formed mostly by the acrylate pathway, was the predominant product with lactate alone. Fluorescent in situ hybridization demonstrated that populations of Bifidobacterium spp., major lactate producers, increased approximately 10-fold in incubations with polysaccharides. Populations of Eubacterium hallii, a lactate-utilizing butyrate-producing bacterium, increased 100-fold at pH 5.9 and 6.4. These experiments suggest that lactate is rapidly converted to acetate, butyrate, and propionate by the human intestinal microbiota at pH values as low as 5.9, but at pH 5.2 reduced utilization occurs while production is maintained, resulting in lactate accumulation.
Figures
References
-
- Al Jassim, R. A. M., P. T. Scott, A. L. Trebbin, D. Trott, and C. C. Pollitt. 2005. The genetic diversity of lactic acid producing bacteria in the equine gastrointestinal tract. FEMS Microbiol. Lett. 248:75-81. - PubMed
-
- Avivi-Green, C., S. Polak-Charcon, Z. Madar, and B. Schwartz. 2000. Apoptosis cascade proteins are regulated in vivo by high intracolonic butyrate concentration: correlation with colon cancer inhibition. Oncol. Res. 12:83-95. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
