

NMR structure of a kissing complex formed between the TAR RNA element of HIV-1 and a LNA-modified aptamer
- PMID: 17768146
- PMCID: PMC2094059
- DOI: 10.1093/nar/gkm655
NMR structure of a kissing complex formed between the TAR RNA element of HIV-1 and a LNA-modified aptamer
Abstract
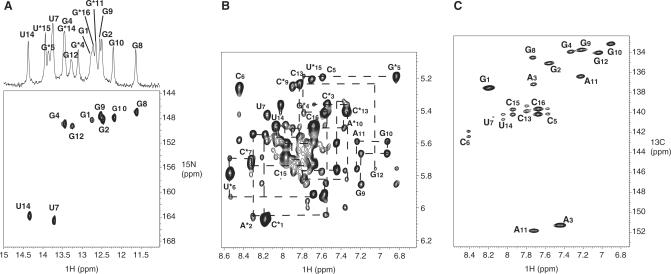
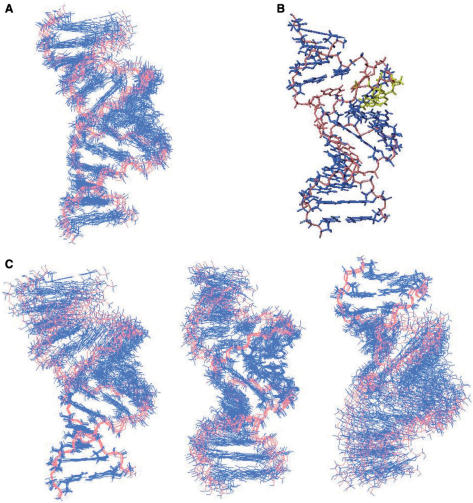
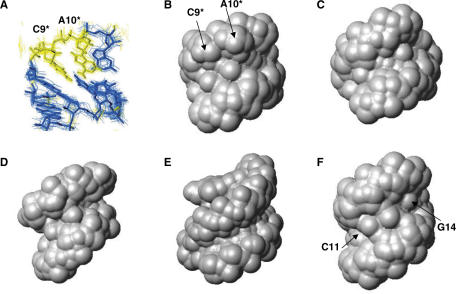
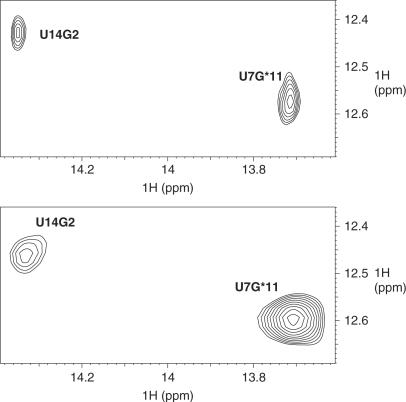
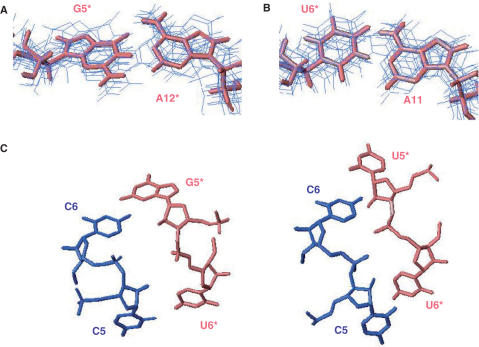
The trans-activating responsive (TAR) RNA element located in the 5' untranslated region of the HIV-1 genome is a 57-nt imperfect stem-loop essential for the viral replication. TAR regulates transcription by interacting with both viral and cellular proteins. RNA hairpin aptamers specific for TAR were previously identified by in vitro selection [Ducongé,F. and Toulmé,J.J. (1999) In vitro selection identifies key determinants for loop-loop interactions: RNA aptamers selective for the TAR RNA element of HIV-1. RNA, 5, 1605-1614]. These aptamers display a 5'-GUCCCAGA-3' consensus apical loop, partially complementary to the TAR one, leading to the formation of a TAR-aptamer kissing complex. The conserved GA combination (underlined in the consensus sequence) has been shown to be crucial for the formation of a highly stable complex. To improve the nuclease resistance of the aptamer and to increase its affinity for TAR, locked nucleic acid (LNA) nucleotides were introduced in the aptamer apical loop. LNA are nucleic acids analogues that contain a 2'-O,4'-C methylene linkage and that raise the thermostablity of duplexes. We solved the NMR solution structure of the TAR-LNA-modified aptamer kissing complex. Structural analysis revealed the formation of a non-canonical G*A pair leading to increased stacking at the stem-loop junction. Our data also showed that the introduction of LNA residues provides an enhanced stability while maintaining a normal Watson-Crick base pairing with a loop-loop conformation close to an A-type.
Figures

Similar articles
-
LNA derivatives of a kissing aptamer targeted to the trans-activating responsive RNA element of HIV-1.Blood Cells Mol Dis. 2007 May-Jun;38(3):204-9. doi: 10.1016/j.bcmd.2006.11.008. Epub 2007 Feb 14. Blood Cells Mol Dis. 2007. PMID: 17300966
-
Aptamers targeted to an RNA hairpin show improved specificity compared to that of complementary oligonucleotides.Biochemistry. 2006 Oct 3;45(39):12076-82. doi: 10.1021/bi0606344. Biochemistry. 2006. PMID: 17002307
-
In vitro selection of RNA aptamers derived from a genomic human library against the TAR RNA element of HIV-1.Biochemistry. 2009 Jul 7;48(26):6278-84. doi: 10.1021/bi802373d. Biochemistry. 2009. PMID: 19496624
-
Regulating eukaryotic gene expression with aptamers.FEBS Lett. 2004 Jun 1;567(1):55-62. doi: 10.1016/j.febslet.2004.03.111. FEBS Lett. 2004. PMID: 15165893 Review.
-
Modulation of RNA function by oligonucleotides recognizing RNA structure.Prog Nucleic Acid Res Mol Biol. 2001;69:1-46. doi: 10.1016/s0079-6603(01)69043-3. Prog Nucleic Acid Res Mol Biol. 2001. PMID: 11550792 Review.
Cited by
-
Crystallographic characterization of CCG repeats.Nucleic Acids Res. 2012 Sep;40(16):8155-62. doi: 10.1093/nar/gks557. Epub 2012 Jun 19. Nucleic Acids Res. 2012. PMID: 22718980 Free PMC article.
-
Non-natural nucleic acids for synthetic biology.Curr Opin Chem Biol. 2009 Dec;13(5-6):687-96. doi: 10.1016/j.cbpa.2009.09.030. Epub 2009 Oct 29. Curr Opin Chem Biol. 2009. PMID: 19879178 Free PMC article. Review.
-
Modelling aptamers with nucleic acid mimics (NAM): From sequence to three-dimensional docking.PLoS One. 2022 Mar 23;17(3):e0264701. doi: 10.1371/journal.pone.0264701. eCollection 2022. PLoS One. 2022. PMID: 35320268 Free PMC article.
-
Exploring TAR-RNA aptamer loop-loop interaction by X-ray crystallography, UV spectroscopy and surface plasmon resonance.Nucleic Acids Res. 2008 Dec;36(22):7146-56. doi: 10.1093/nar/gkn831. Epub 2008 Nov 7. Nucleic Acids Res. 2008. PMID: 18996893 Free PMC article.
-
Characterizing complex dynamics in the transactivation response element apical loop and motional correlations with the bulge by NMR, molecular dynamics, and mutagenesis.Biophys J. 2008 Oct;95(8):3906-15. doi: 10.1529/biophysj.108.140285. Epub 2008 Jul 11. Biophys J. 2008. PMID: 18621815 Free PMC article.
References
-
- Muesing MA, Smith DH, Capon DJ. Regulation of mRNA accumulation by a human immunodeficiency virus trans-activator protein. Cell. 1987;48:691–701. - PubMed
-
- Isel C, Karn J. Direct evidence that HIV-1 Tat stimulates RNA polymerase II carboxyl-terminal domain hyperphosphorylation during transcriptional elongation. J. Mol. Biol. 1999;290:929–941. - PubMed
-
- Berkhout B. Multiple biological roles associated with the repeat (R) region of the HIV-1 RNA genome. Adv. Pharmacol. 2000;48:29–73. - PubMed
-
- Berkhout B, Silverman RH, Jeang KT. Tat trans-activates the human immunodeficiency virus through a nascent RNA target. Cell. 1989;59:273–282. - PubMed
-
- Karn J. Tackling Tat. J. Mol. Biol. 1999;293:235–254. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
