

Induction of the CTLA-4 gene in human lymphocytes is dependent on NFAT binding the proximal promoter
- PMID: 17785820
- PMCID: PMC4290020
- DOI: 10.4049/jimmunol.179.6.3831
Induction of the CTLA-4 gene in human lymphocytes is dependent on NFAT binding the proximal promoter
Abstract
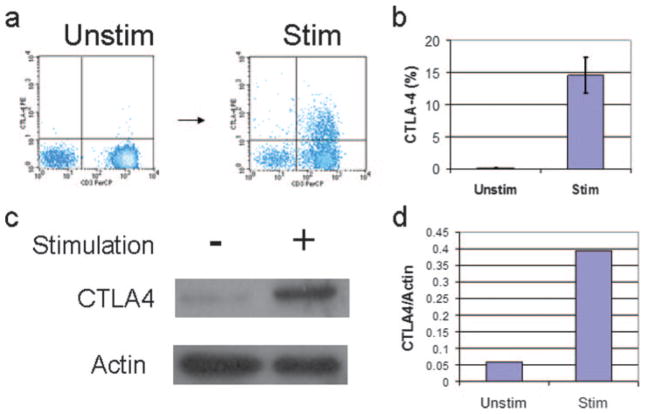
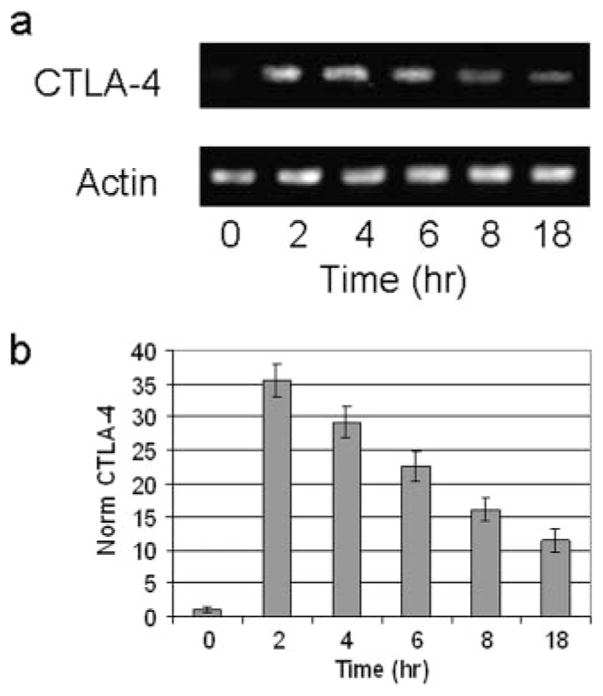
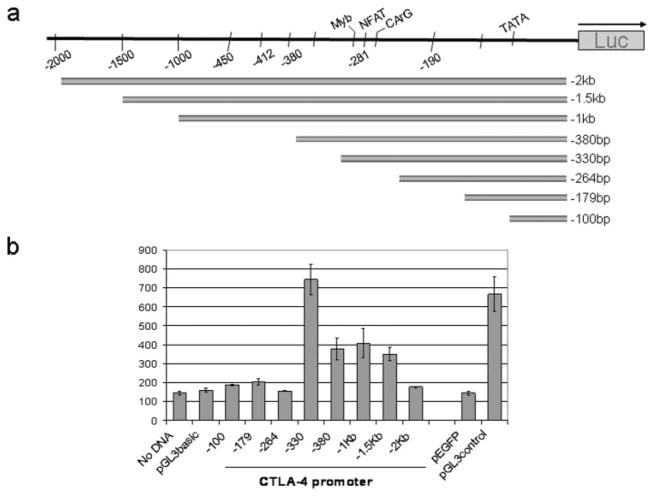
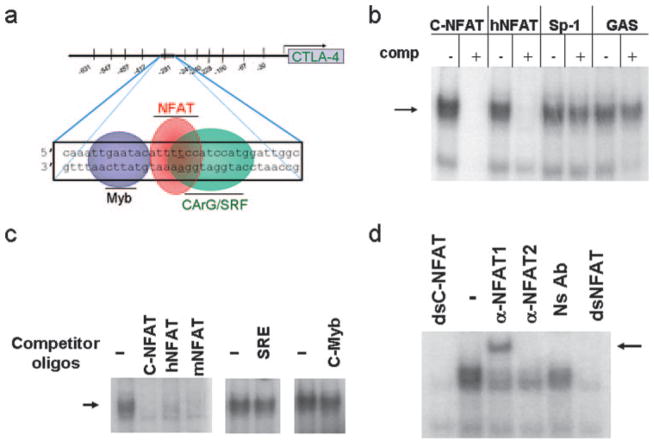
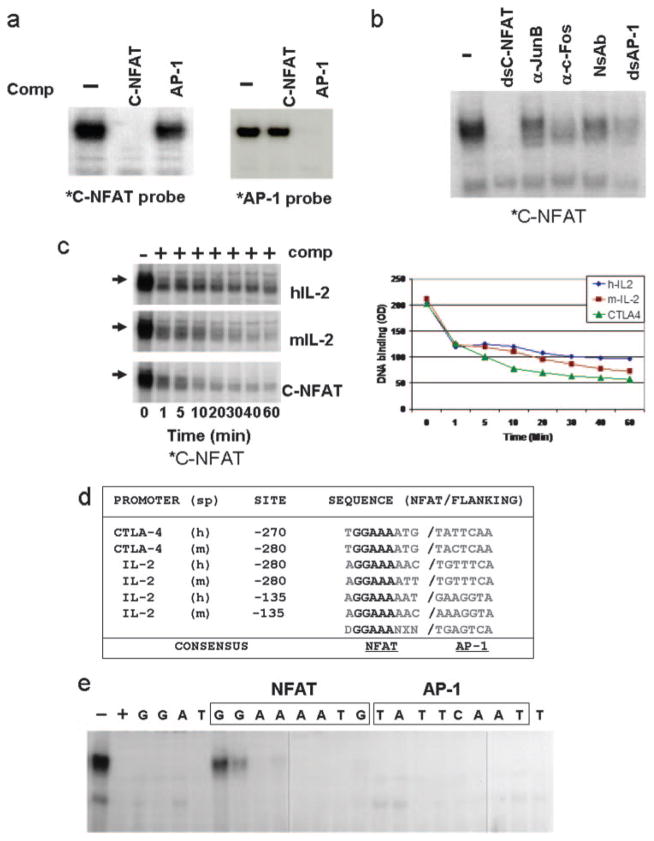
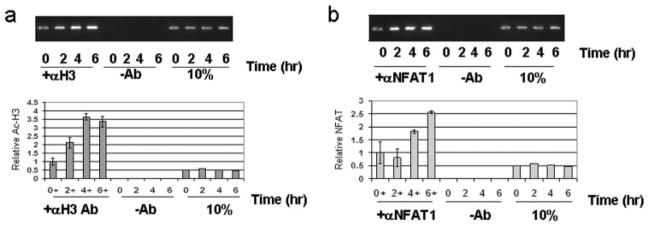
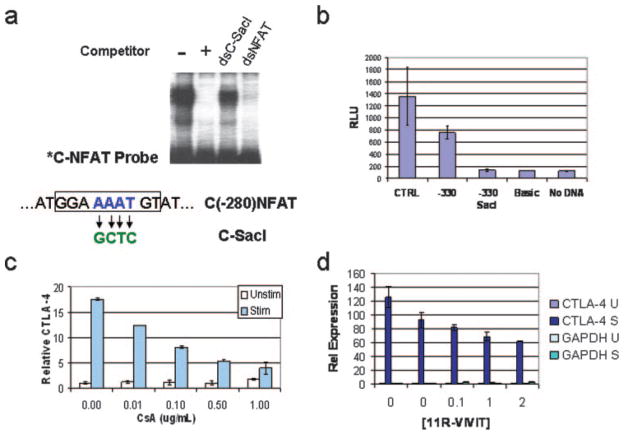
CTLA-4 is a member of the costimulatory family, has homology to CD28, and binds the B7 family of ligands. Unlike CD28, CTLA-4 ligation transmits a negative signal in T cells. CTLA-4 expression, while inducible in most T cells, is expressed constitutively on T cells with a regulatory phenotype. The mechanism controlling CTLA-4 expression in human T cells is poorly characterized, thus we sought to better understand the mechanism of activation of the CTLA-4 gene. By cloning the 5' upstream promoter and creating promoter-deletion reporter constructs, we show that the proximal promoter is critical for activating the CTLA-4 gene. Within this region, we identify a NFAT consensus sequence that binds NFAT with high affinity that differs from other NFAT sequences and does not recruit AP-1. Analysis of the chromatin proteins in the native CTLA-4 gene shows that this promoter region becomes associated with acetylated histones by chromatin immunoprecipitation assays. In addition, NFAT1 binds to the promoter of the CTLA-4 gene after stimulation by chromatin immunoprecipitation. The functional requirement of the NFAT site for CTLA-4 transcription was demonstrated by mutations in the NFAT site that abolished the activity of the promoter. Furthermore, inhibitors of NFAT suppressed CTLA-4 gene expression, indicating that NFAT plays a critical role in regulating the induction of the CTLA-4 gene in lymphocytes. The identification of NFAT as a critical regulator of the CTLA-4 gene suggests that targeting NFAT function may lead to novel approaches to modulate the CTLA-4 gene to control the immune response.
Conflict of interest statement
The authors have no financial conflict of interest.
Figures
References
-
- Greenfield EA, Nguyen KA, Kuchroo VK. CD28/B7 costimulation: a review. Crit Rev Immunol. 1998;18:389–418. - PubMed
-
- Slavik JM, Hutchcroft JE, Bierer BE. CD28/CTLA-4 and CD80/CD86 families: signaling and function. Immunol Res. 1999;19:1–24. - PubMed
-
- Yu X, Fournier S, Allison JP, Sharpe AH, Hodes RJ. The role of B7 costimulation in CD4/CD8 T cell homeostasis. J Immunol. 2000;164:3543–3553. - PubMed
-
- Chambers CA, Allison JP. Costimulatory regulation of T cell function. Curr Opin Cell Biol. 1999;11:203–210. - PubMed
-
- Teft WA, Kirchhof MG, Madrenas J. A molecular perspective of CTLA-4 function. Annu Rev Immunol. 2006;24:65–97. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
