

Histone deacetylase degradation and MEF2 activation promote the formation of slow-twitch myofibers
- PMID: 17786239
- PMCID: PMC1957540
- DOI: 10.1172/JCI31960
Histone deacetylase degradation and MEF2 activation promote the formation of slow-twitch myofibers
Abstract
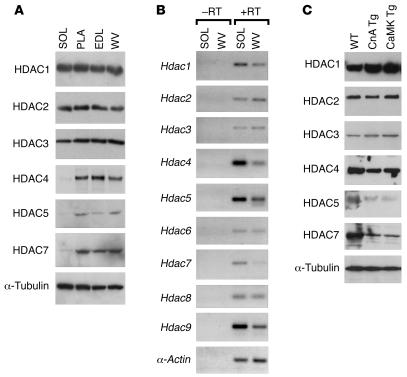
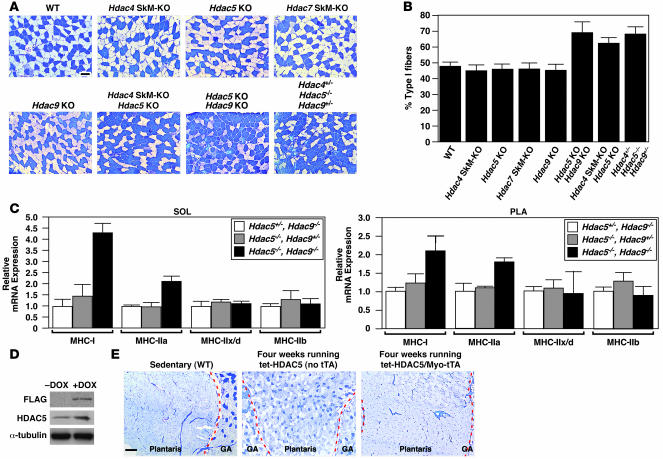
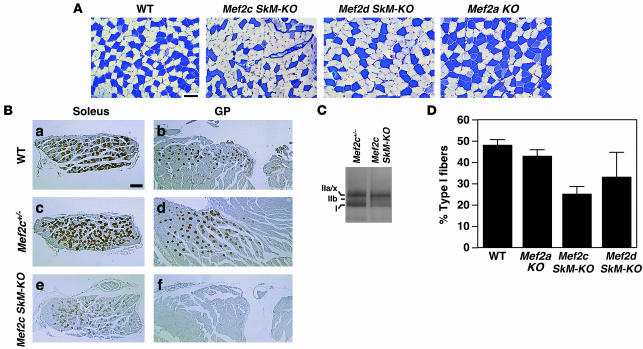
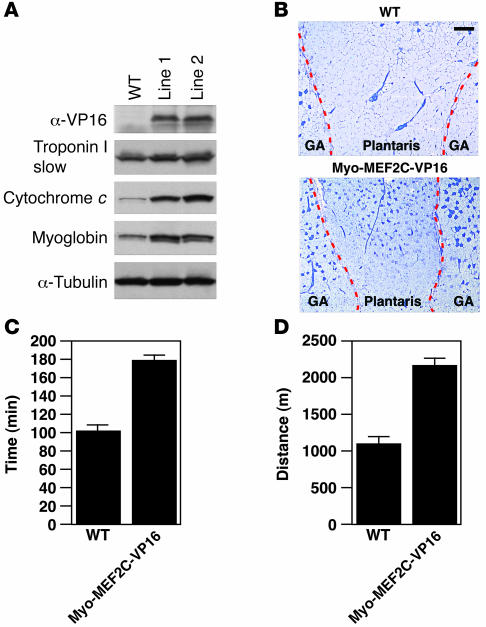
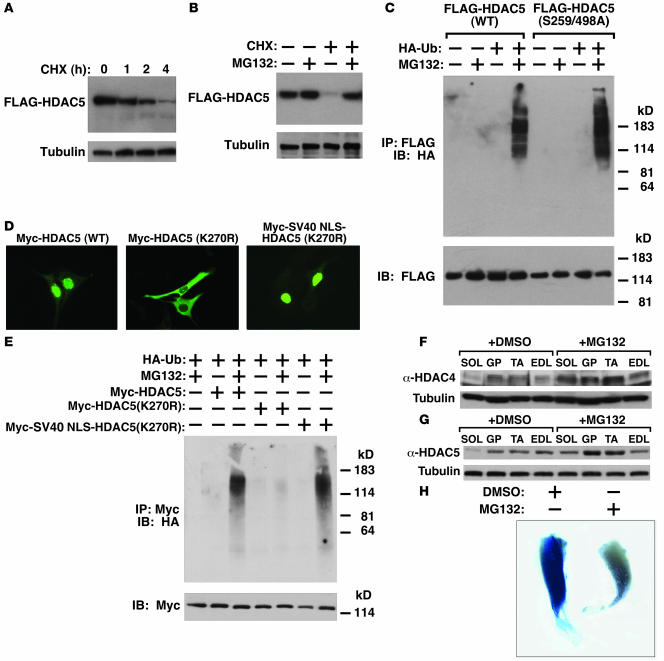
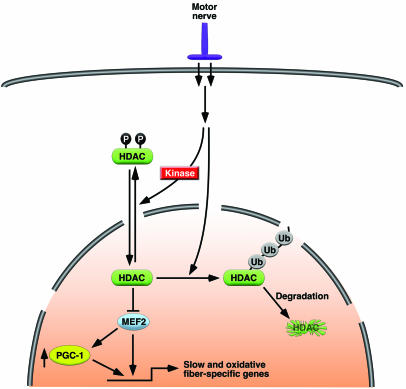
Skeletal muscle is composed of heterogeneous myofibers with distinctive rates of contraction, metabolic properties, and susceptibility to fatigue. We show that class II histone deacetylase (HDAC) proteins, which function as transcriptional repressors of the myocyte enhancer factor 2 (MEF2) transcription factor, fail to accumulate in the soleus, a slow muscle, compared with fast muscles (e.g., white vastus lateralis). Accordingly, pharmacological blockade of proteasome function specifically increases expression of class II HDAC proteins in the soleus in vivo. Using gain- and loss-of-function approaches in mice, we discovered that class II HDAC proteins suppress the formation of slow twitch, oxidative myofibers through the repression of MEF2 activity. Conversely, expression of a hyperactive form of MEF2 in skeletal muscle of transgenic mice promotes the formation of slow fibers and enhances running endurance, enabling mice to run almost twice the distance of WT littermates. Thus, the selective degradation of class II HDACs in slow skeletal muscle provides a mechanism for enhancing physical performance and resistance to fatigue by augmenting the transcriptional activity of MEF2. These findings provide what we believe are new insights into the molecular basis of skeletal muscle function and have important implications for possible therapeutic interventions into muscular diseases.
Figures
Comment in
-
Two tales concerning skeletal muscle.J Clin Invest. 2007 Sep;117(9):2388-91. doi: 10.1172/JCI33379. J Clin Invest. 2007. PMID: 17786235 Free PMC article.
References
-
- Pette D., Staron R.S. Myosin isoforms, muscle fiber types, and transitions. Microsc. Res. Tech. 2000;50:500–509. - PubMed
-
- Schiaffino S., Reggiani C. Molecular diversity of myofibrillar proteins: gene regulation and functional significance. Physiol. Rev. 1996;76:371–423. - PubMed
-
- Baldwin K.M., Haddad F. Effects of different activity and inactivity paradigms on myosin heavy chain gene expression in striated muscle. J. Appl. Physiol. 2001;90:345–357. - PubMed
-
- Sugiura T., Miyata H., Kawai Y., Matoba H., Murakami N. Changes in myosin heavy chain isoform expression of overloaded rat skeletal muscles. Int. J. Biochem. 1993;25:1609–1613. - PubMed
-
- Talmadge R.J. Myosin heavy chain isoform expression following reduced neuromuscular activity: potential regulatory mechanisms. Muscle Nerve. 2000;23:661–679. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
