

Exonuclease-1 deletion impairs DNA damage signaling and prolongs lifespan of telomere-dysfunctional mice
- PMID: 17803909
- PMCID: PMC2658812
- DOI: 10.1016/j.cell.2007.08.029
Exonuclease-1 deletion impairs DNA damage signaling and prolongs lifespan of telomere-dysfunctional mice
Erratum in
- Cell. 2007 Oct 5;131(1):190. Kuhnel, Florian [added]
Abstract
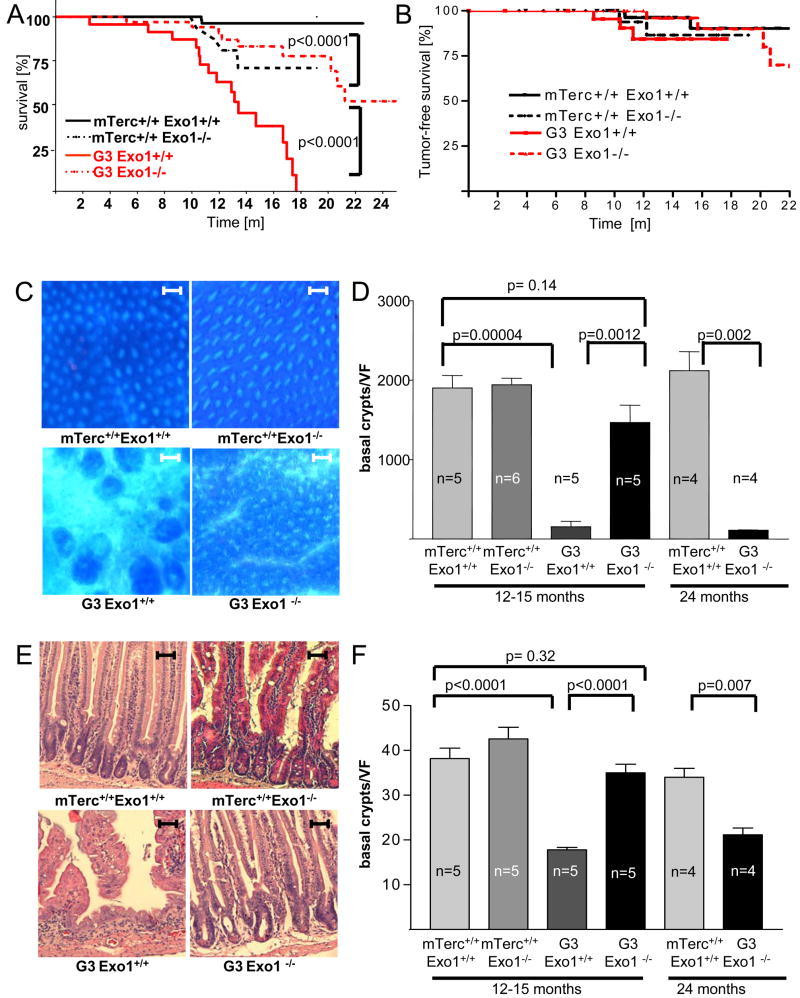
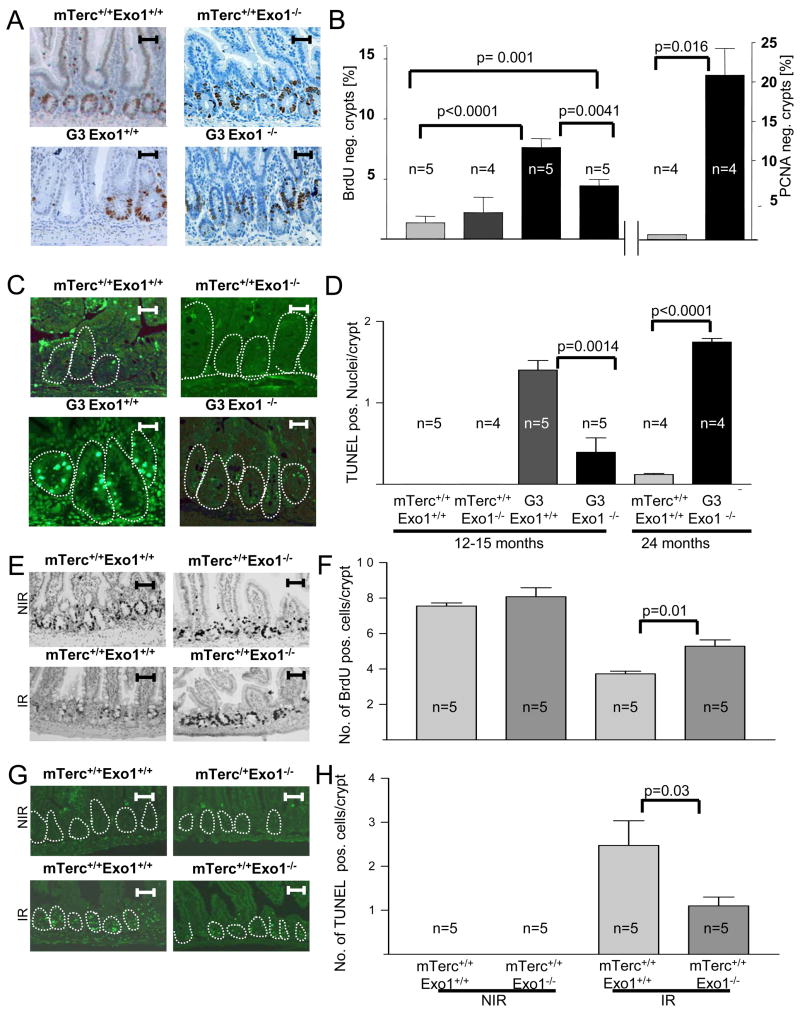
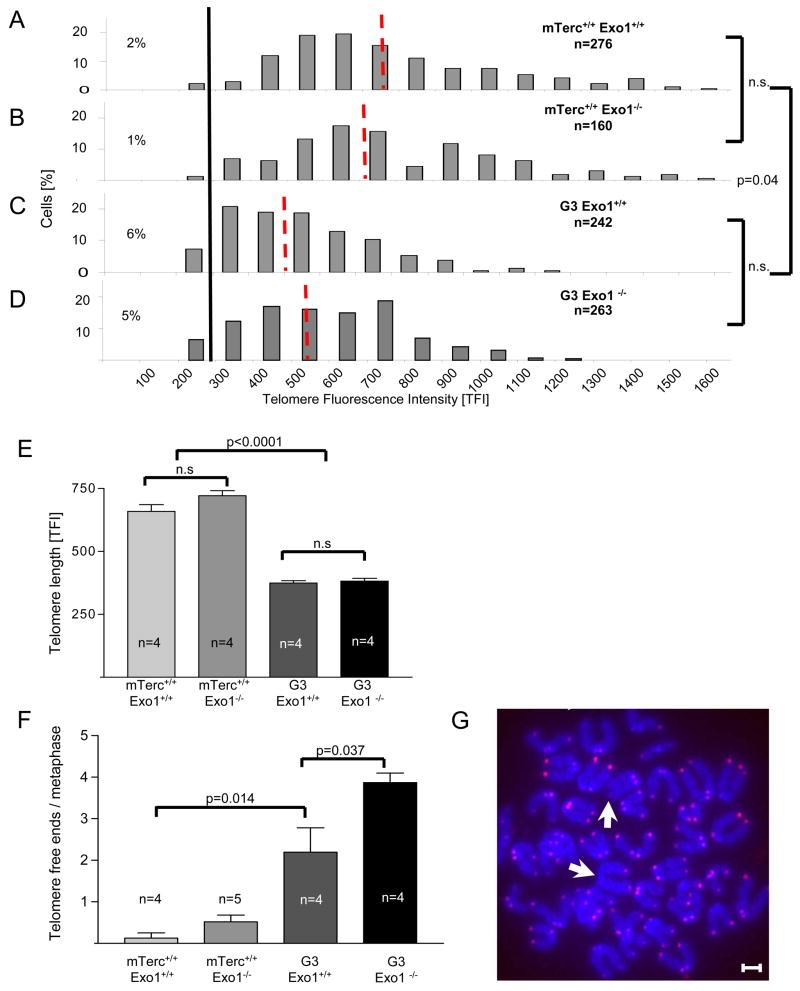
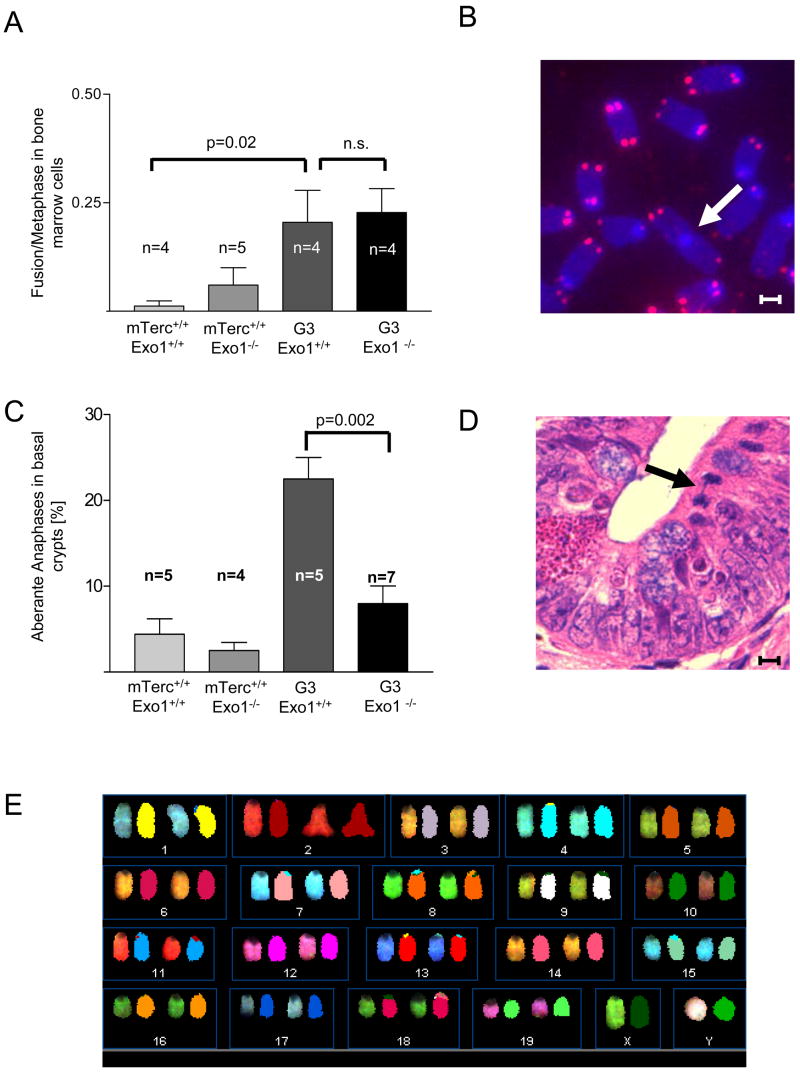
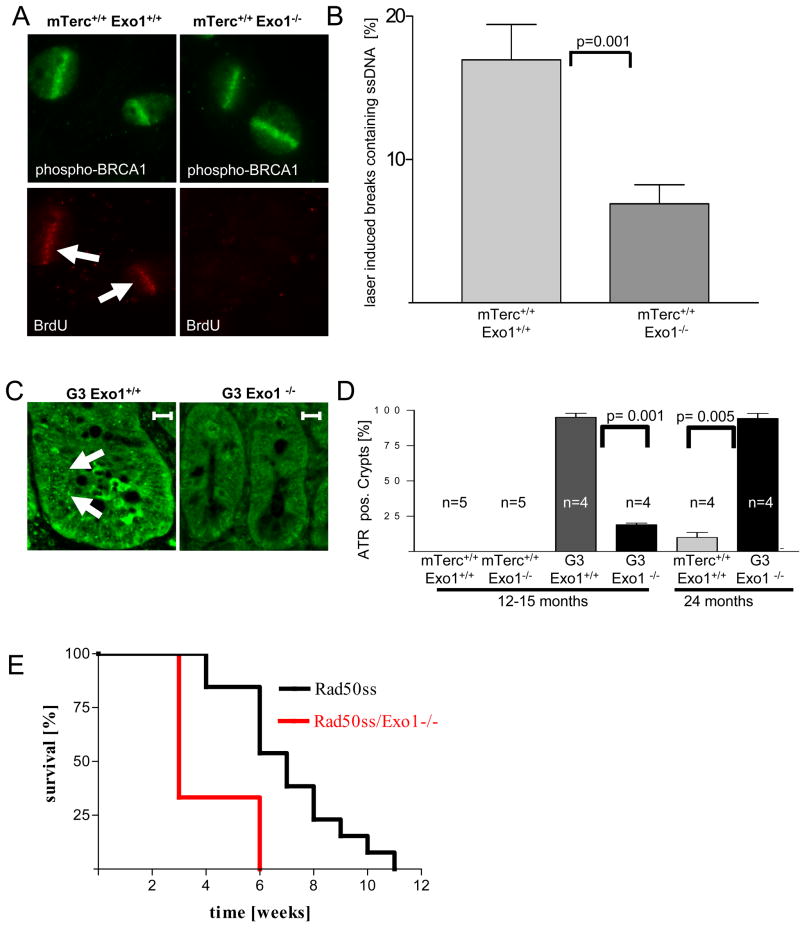
Exonuclease-1 (EXO1) mediates checkpoint induction in response to telomere dysfunction in yeast, but it is unknown whether EXO1 has similar functions in mammalian cells. Here we show that deletion of the nuclease domain of Exo1 reduces accumulation of DNA damage and DNA damage signal induction in telomere-dysfunctional mice. Exo1 deletion improved organ maintenance and lifespan of telomere-dysfunctional mice but did not increase chromosomal instability or cancer formation. Deletion of Exo1 also ameliorated the induction of DNA damage checkpoints in response to gamma-irradiation and conferred cellular resistance to 6-thioguanine-induced DNA damage. Exo1 deletion impaired upstream induction of DNA damage responses by reducing ssDNA formation and the recruitment of Replication Protein A (RPA) and ATR at DNA breaks. Together, these studies provide evidence that EXO1 contributes to DNA damage signal induction in mammalian cells, and deletion of Exo1 can prolong survival in the context of telomere dysfunction.
Figures


References
-
- Armanios MY, Chen JJ, Cogan JD, Alder JK, Ingersoll RG, Markin C, Lawson WE, Xie M, Vulto I, Phillips JA, 3rd, et al. Telomerase mutations in families with idiopathic pulmonary fibrosis. N Engl J Med. 2007;356:1317–1326. - PubMed
-
- Artandi SE, Chang S, Lee SL, Alson S, Gottlieb GJ, Chin L, DePinho RA. Telomere dysfunction promotes non-reciprocal translocations and epithelial cancers in mice. Nature. 2000;406:641–645. - PubMed
-
- Ball SE, Gibson FM, Rizzo S, Tooze JA, Marsh JC, Gordon-Smith EC. Progressive telomere shortening in aplastic anemia. Blood. 1998;91:3582–3592. - PubMed
-
- Blackburn EH. Switching and signaling at the telomere. Cell. 2001;106:661–673. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
