

NF-kappaB is a negative regulator of IL-1beta secretion as revealed by genetic and pharmacological inhibition of IKKbeta
- PMID: 17803913
- PMCID: PMC2134986
- DOI: 10.1016/j.cell.2007.07.009
NF-kappaB is a negative regulator of IL-1beta secretion as revealed by genetic and pharmacological inhibition of IKKbeta
Abstract
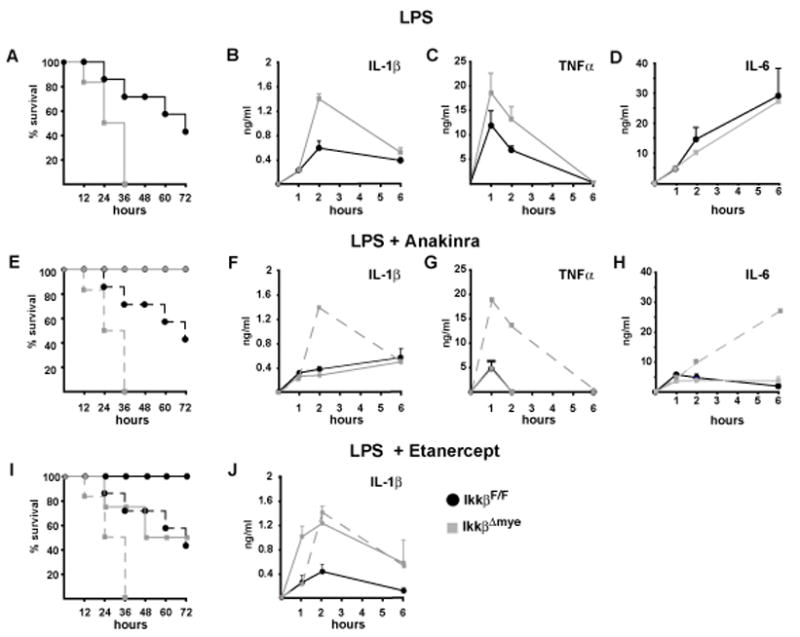
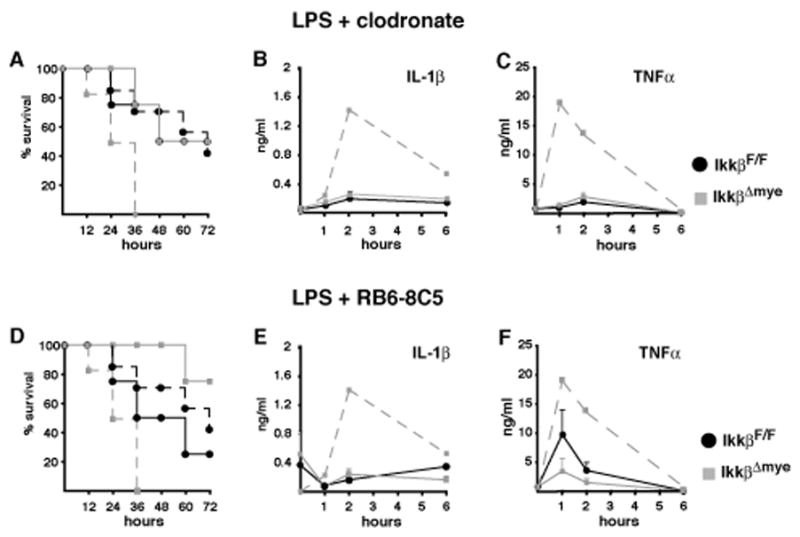
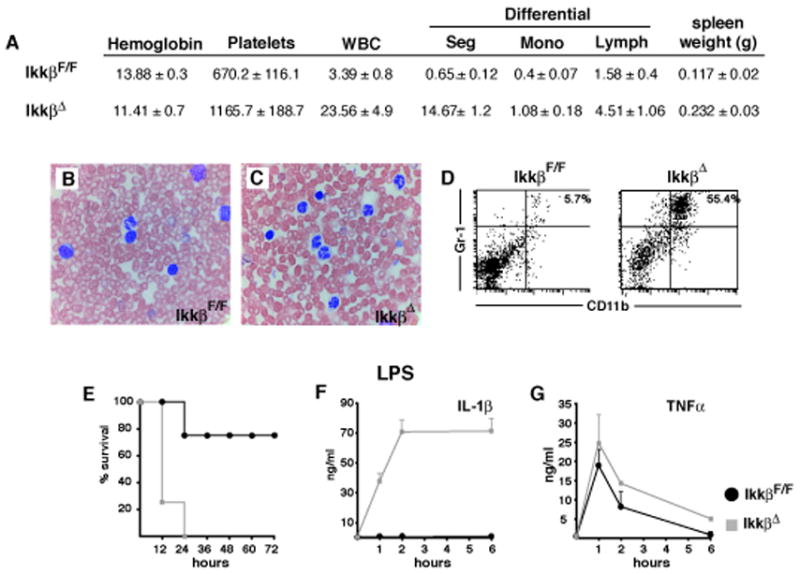
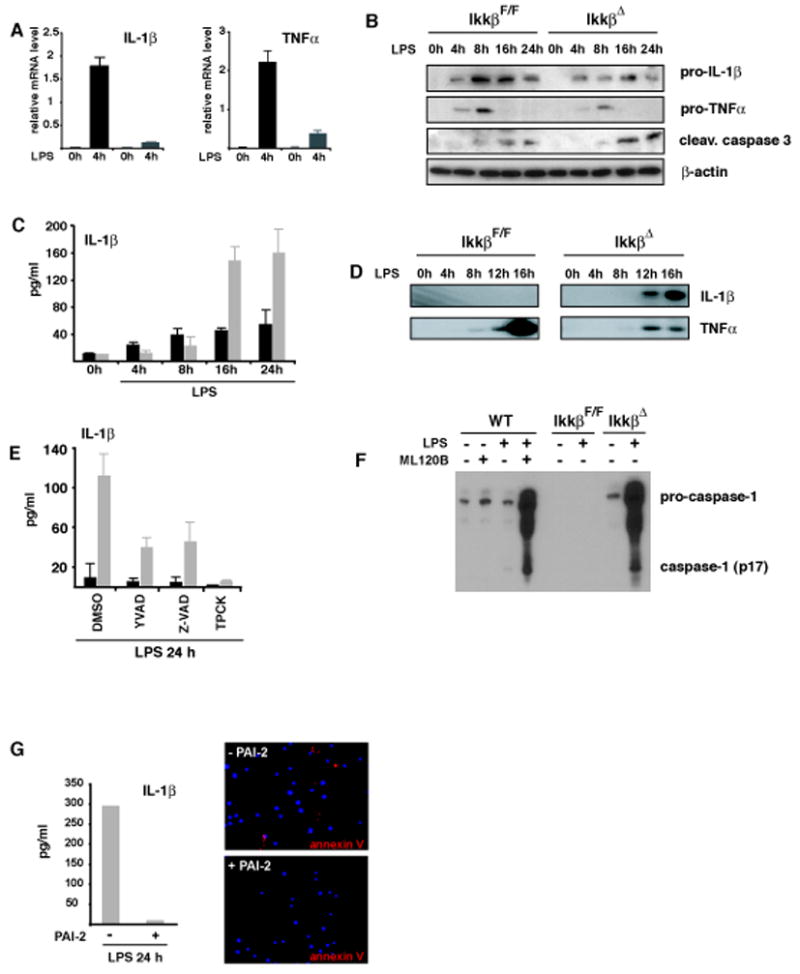
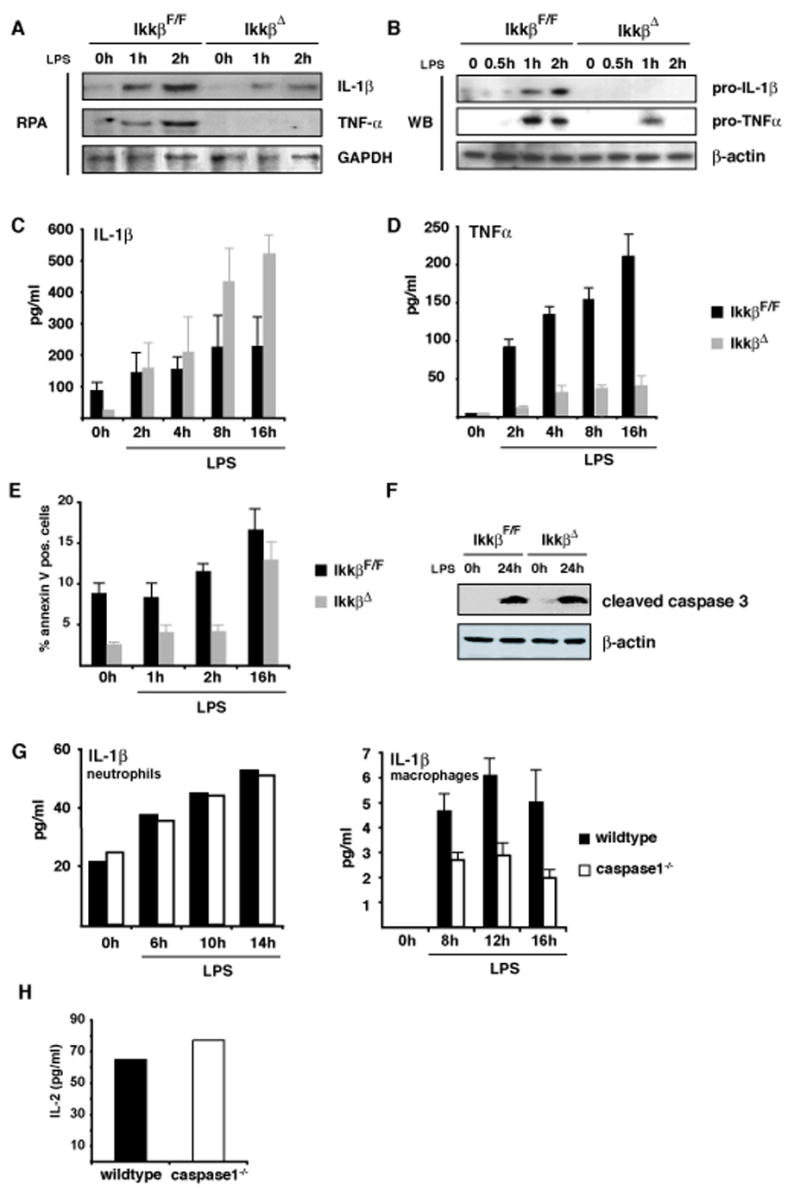
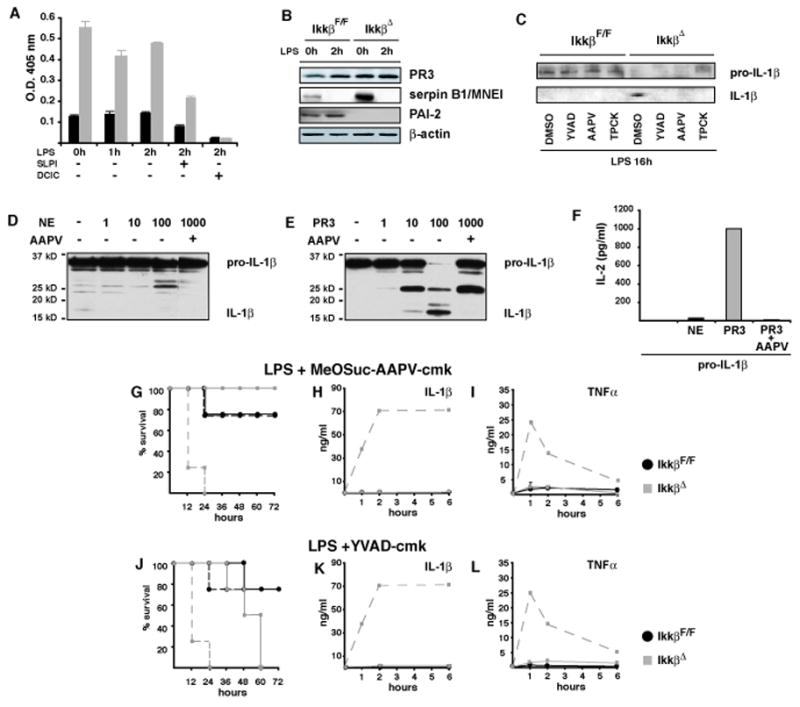
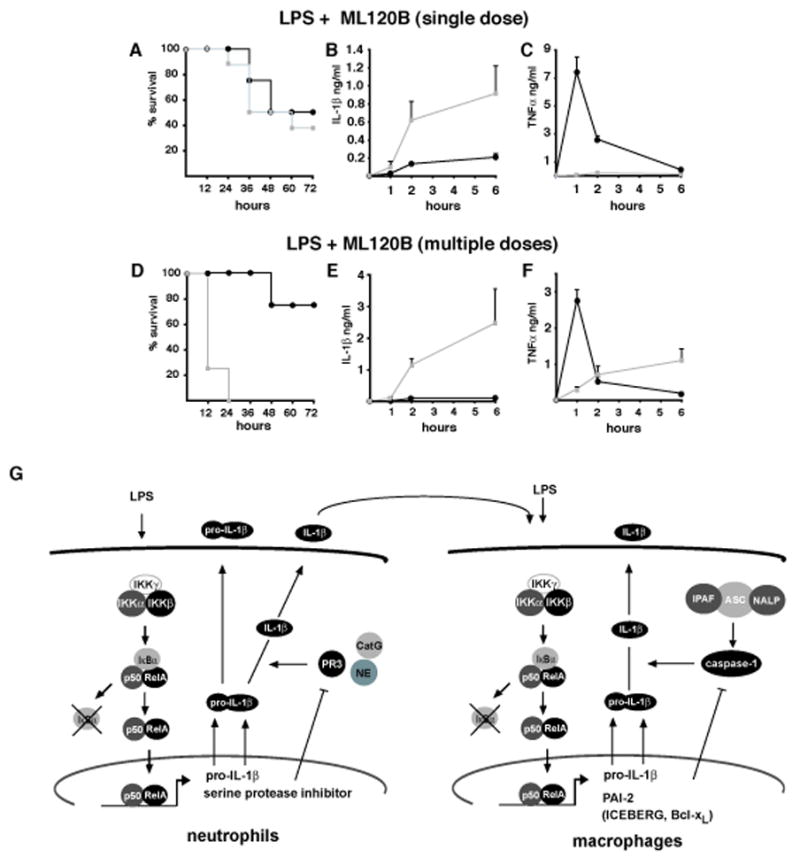
IKKbeta-dependent NF-kappaB activation plays a key role in innate immunity and inflammation, and inhibition of IKKbeta has been considered as a likely anti-inflammatory therapy. Surprisingly, however, mice with a targeted IKKbeta deletion in myeloid cells are more susceptible to endotoxin-induced shock than control mice. Increased endotoxin susceptibility is associated with elevated plasma IL-1beta as a result of increased pro-IL-1beta processing, which was also seen upon bacterial infection. In macrophages enhanced pro-IL-1beta processing depends on caspase-1, whose activation is inhibited by NF-kappaB-dependent gene products. In neutrophils, however, IL-1beta secretion is caspase-1 independent and depends on serine proteases, whose activity is also inhibited by NF-kappaB gene products. Prolonged pharmacologic inhibition of IKKbeta also augments IL-1beta secretion upon endotoxin challenge. These results unravel an unanticipated role for IKKbeta-dependent NF-kappaB signaling in the negative control of IL-1beta production and highlight potential complications of long-term IKKbeta inhibition.
Figures
References
-
- Arkan MC, Hevener AL, Greten FR, Maeda S, Li ZW, Long JM, Wynshaw-Boris A, Poli G, Olefsky J, Karin M. IKK-beta links inflammation to obesity-induced insulin resistance. Nat Med. 2005;11:191–198. - PubMed
-
- Black RA, Kronheim SR, Cantrell M, Deeley MC, March CJ, Prickett KS, Wignall J, Conlon PJ, Cosman D, Hopp TP, et al. Generation of biologically active interleukin-1 beta by proteolytic cleavage of the inactive precursor. J Biol Chem. 1988;263:9437–9442. - PubMed
-
- Bonizzi G, Karin M. The two NF-kappaB activation pathways and their role in innate and adaptive immunity. Trends Immunol. 2004;25:280–288. - PubMed
-
- Bruey JM, Bruey-Sedano N, Luciano F, Zhai D, Balpai R, Xu C, Kress CL, Bailly-Maitre B, Li X, Osterman A, et al. Bcl-2 and Bcl-XL regulate proinflammatory caspase-1 activation by interaction with NALP1. Cell. 2007;129:45–56. - PubMed
-
- Chen LW, Egan L, Li ZW, Greten FR, Kagnoff MF, Karin M. The two faces of IKK and NF-kappaB inhibition: prevention of systemic inflammation but increased local injury following intestinal ischemia-reperfusion. Nat Med. 2003;9:575–581. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
