

Comparative Study
doi: 10.1016/j.molcel.2007.06.028.
A molecular brake in the kinase hinge region regulates the activity of receptor tyrosine kinases
Affiliations
- PMID: 17803937
- PMCID: PMC2094128
- DOI: 10.1016/j.molcel.2007.06.028
Item in Clipboard
Comparative Study
A molecular brake in the kinase hinge region regulates the activity of receptor tyrosine kinases
Mol Cell.
.
Abstract
Activating mutations in the tyrosine kinase domain of receptor tyrosine kinases (RTKs) cause cancer and skeletal disorders. Comparison of the crystal structures of unphosphorylated and phosphorylated wild-type FGFR2 kinase domains with those of seven unphosphorylated pathogenic mutants reveals an autoinhibitory "molecular brake" mediated by a triad of residues in the kinase hinge region of all FGFRs. Structural analysis shows that many other RTKs, including PDGFRs, VEGFRs, KIT, CSF1R, FLT3, TEK, and TIE, are also subject to regulation by this brake. Pathogenic mutations activate FGFRs and other RTKs by disengaging the brake either directly or indirectly.
Figures
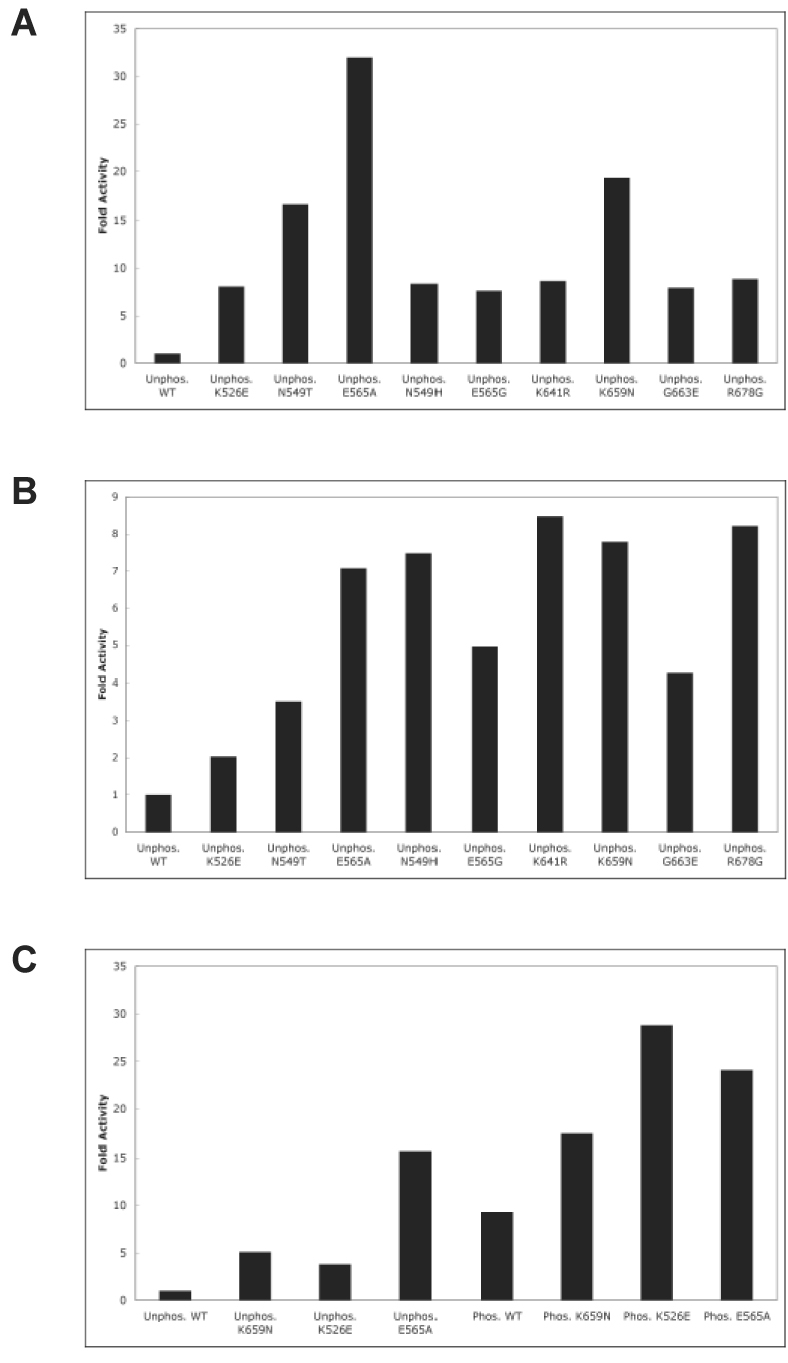
Pathogenic mutations activate FGFR2K constitutively in the absence of A-loop tyrosine phosphorylation. (A) Autophosphorylation assays: unphosphorylated wild-type or mutant FGFR2Ks were incubated with 1 mM ATP and autophosphorylation rates were determined using a continuous spectrophotometric assay. (B) Peptide phosphorylation assays: unphosphorylated wild-type or mutant FGFR2Ks were incubated with 100 µM ATP and 50 µM peptide substrate (KKEEEEYMMMMG). Rates of peptide phosphorylation were determined using the spectrophotometric assay. (C) The effect of A-loop phosphorylation on kinase activity of wild-type and mutant kinases. Unphosphorylated and A-loop tyrosine phosphorylated wild-type, K526E, K565 or K659N mutant FGFR2Ks were incubated with 500 µM ATP and 500 µM peptide substrate (KKEEEEYMMMMG). Rates of peptide phosphorylation were determined using the spectrophotometric assay.
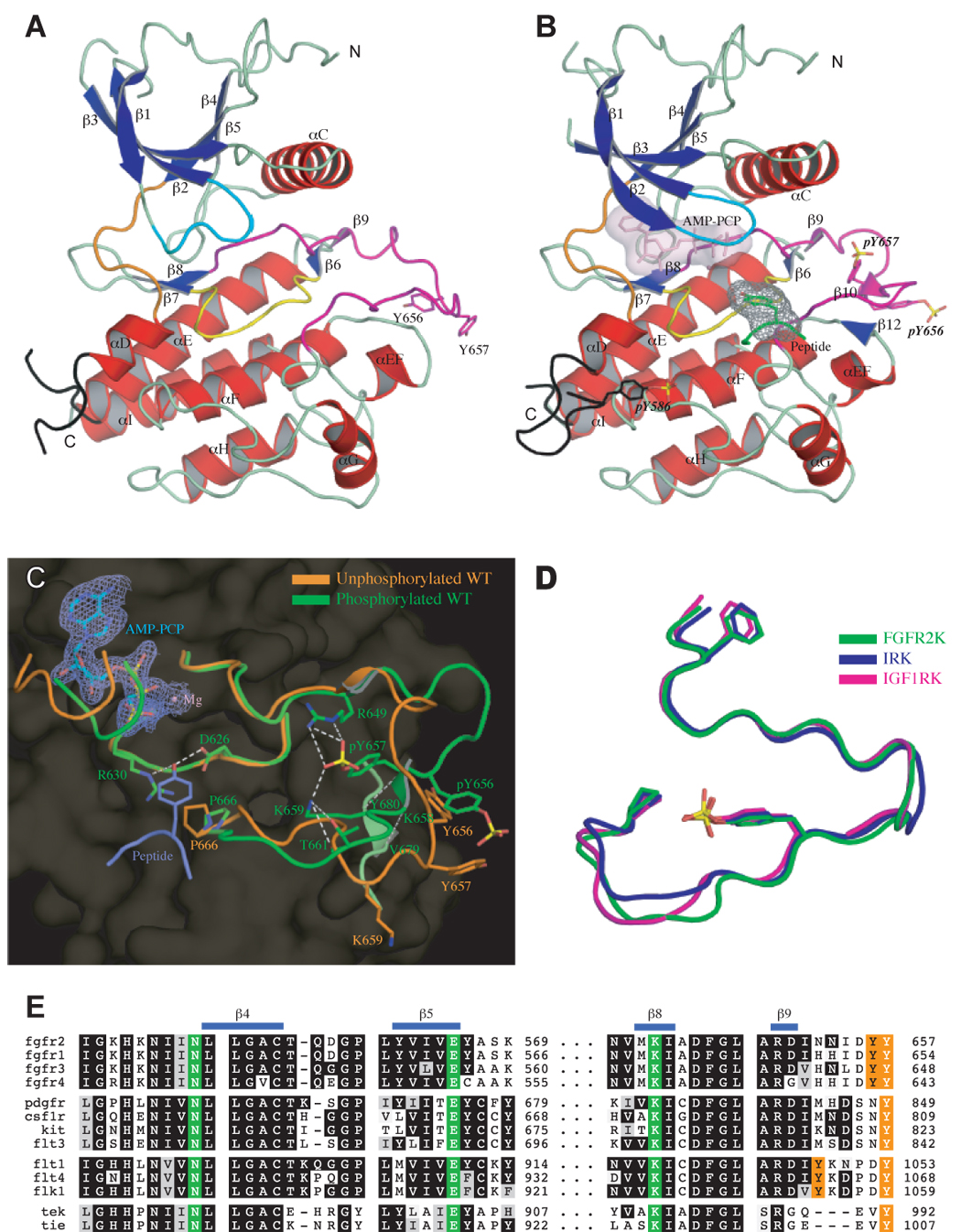
Structural basis for FGFR2K activation by phosphorylation of A-loop tyrosine. (A) and (B) The ribbon diagrams of unphosphorylated and A-loop tyrosine phosphorylated wild-type FGFR2K structures in the same orientation. Strands and helices are colored blue and red, respectively. The A-loop, catalytic loop, nucleotide-binding loop, kinase insert and kinase hinge are colored magenta, yellow, cyan, black, and orange, respectively. In the phosphorylated structure, the ATP analogue (in light pink) and the tyrosine side chain of the peptide substrate (in green) are rendered as sticks. The molecular surfaces of the ATP analogue and the substrate tyrosine are also shown as a solid semi-transparent surface and gray mesh, respectively. (C) Comparison of the A-loop conformations of unphosphorylated (in orange) and A-loop tyrosine phosphorylated wild-type (in green) FGFR2K structures. The ATP analogue (Cα in cyan) and substrate tyrosine (Cα in purple) are shown as sticks. The 2FO-FC electron density map contoured at 1σ for ATP analogue is shown as mesh in slate. Side chains of selected residues from the A-loop and the catalytic loop are shown as sticks. Atom colorings are as follows: oxygens in red, nitrogens in blue, phosphorus in yellow, and carbons are colored according to the kinase molecule to which they belong. Hydrogen bonds are shown as white dashed lines. (D) Comparison of the A-loop conformation of A-loop tyrosine phosphorylated FGFR2K (in green) with those of phosphorylated IRK (PDB ID: 1IR3) (in blue) and IGF1RK (PDB ID: 1K3A) (in magenta). The phosphotyrosine pY657 in FGFR2K fulfills the same function as pY1163 in IRK and pY1133 in IGF1RK. To assist the readers, the side chains of the phenylalanine from the conserved DFG motif at the N-terminus of the A-loop, and of the conserved proline at the C-terminus of the A-loop are also shown as sticks. Coloring scheme is the same as in (C). (E) The triad of residues (in green), which mediate the autoinhibitory molecular brake in FGFR, are also conserved in many other RTKs including PDGFs, CSF1R, KIT, FLT3, VEGFRs, Tie and Tek. Autophosphorylation sites in the A-loop are colored orange.
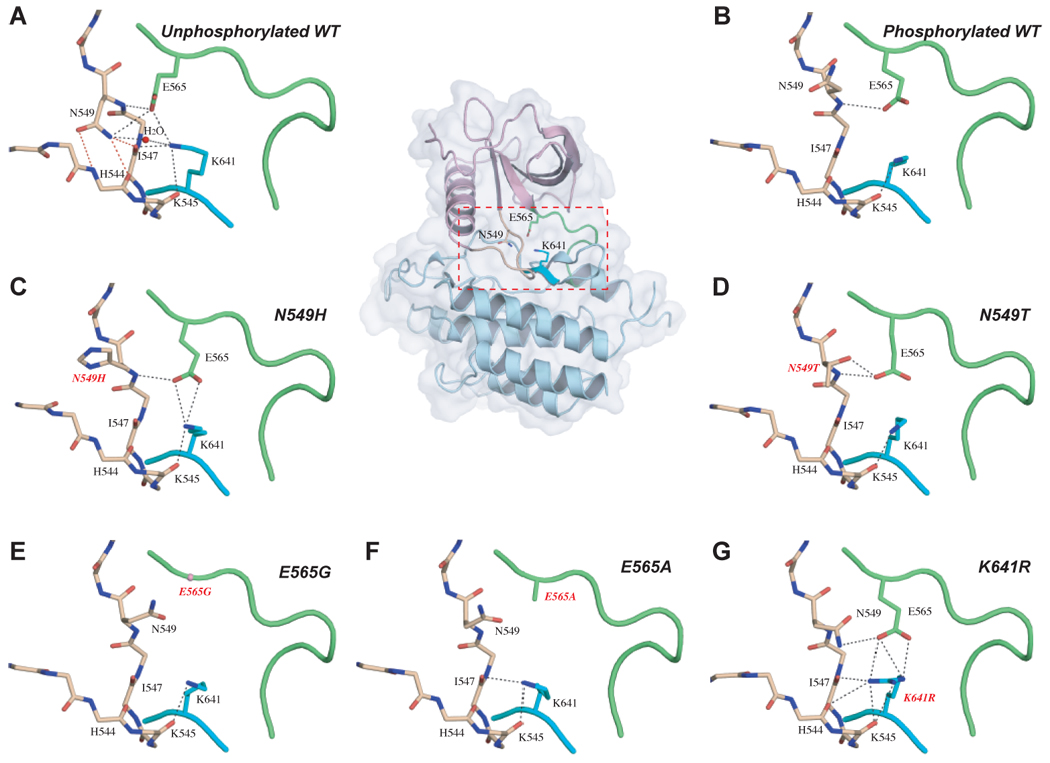
The molecular brake at the kinase hinge region of FGFR2K regulates the kinase activation and is disengaged either by A-loop tyrosine phosphorylation or directly by the pathogenic mutations. (A) In the unphosphorylated wild-type structure, residues N549, E565 and K641 form a network of hydrogen bonds in the kinase hinge region, which serves as a molecular brake to keep the enzyme in an inactive state. (B) The molecular brake is disengaged in the A-loop tyrosine phosphorylated wild-type FGFR2K structure. This molecular brake is also disengaged in the unphosphorylated mutant FGFR2K structures ((C) through (G)). To assist the readers, the whole unphosophorylated wild-type FGFR2K structure is also shown in cartoon and solid semi-transparent surface, and the kinase hinge region is boxed. Atom colorings are as follows: oxygens in red, nitrogens in blue, and carbons are colored according to the kinase region to which they belong. The kinase hinge, the αC-β4 loop (shown in sticks in (A) to (G)), and β8 strand are colored green, wheat and cyan, respectively. The rest of the N-lobe and C-lobe are colored light purple and light blue, respectively. The three critical hydrogen bonds between N549 and the backbone atoms of αC-β4 loop are highlighted by red dashed lines. The remaining hydrogen bonds are shown as black dashed lines.
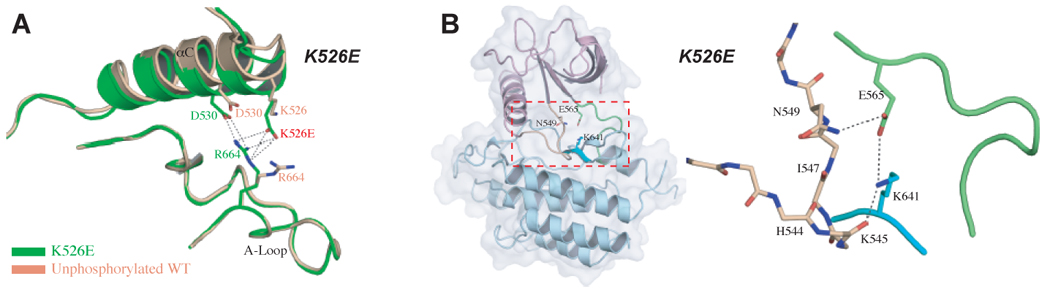
Molecular basis by which the K526E mutation activates FGFR2K in the absence of A-loop tyrosine phosphorylation. (A) The pathogenic K526E mutation (in green) creates new hydrogen bonds between the αC helix and the A-loop which contribute to the inward rotation of the N-lobe. Note that this hydrogen bonding does not occur in the unphosphorylated wild-type FGFR2K structure (in wheat) (B) The molecular brake in the hinge region of the unphosphorylated K526E mutant structure is disengaged indirectly due to the additional hydrogen bonds between the αC helix and the A-loop. To assist the readers, the whole unphosophorylated K526E mutant FGFR2K structure is also shown in cartoon and solid semi-transparent surface, and the kinase hinge region is boxed. Coloring scheme in (B) is as in Fig. 3.
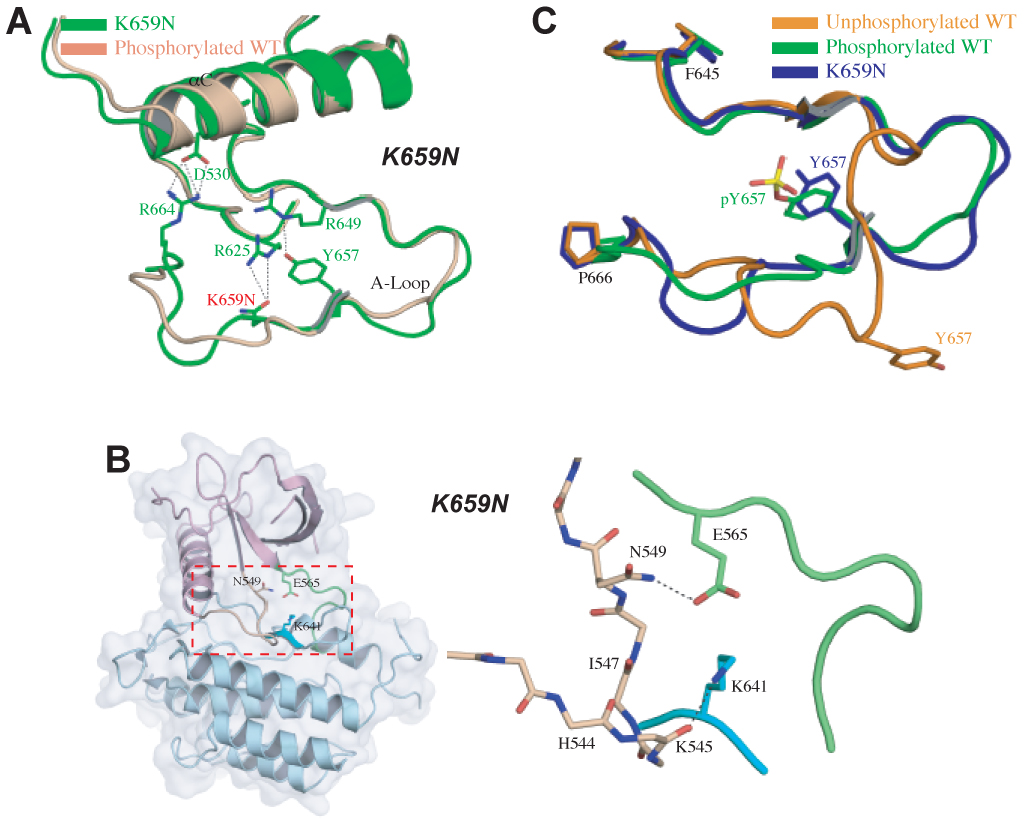
Molecular basis by which the K659N mutation activates FGFR2K in the absence of A-loop tyrosine phosphorylation. (A) Side chain of mutated N659 (in green) in the A-loop hydrogen-bonds with that of R625 in the catalytic loop forcing the A-loop to adopt its active conformation as seen in A-loop tyrosine phosphorylated wild-type FGFR2K structure (in wheat). (B) shows the disengagement of the molecular brake in the hinge region of K659N mutant structure. To assist the readers, the whole unphosophorylated K659E mutant FGFR2K structure is also shown in cartoon and solid semi-transparent surface, and the kinase hinge region is boxed. Coloring scheme in (B) is as in Fig. 3. (C) The K659N mutation drives the A-loop to adopt its active conformation in the absence of A-loop tyrosine phosphorylation. Unphosphorylated wild-type, A-loop phosphorylated wild-type and unphosphorylated K659N mutant FGFR2Ks are shown in orange, green and blue, respectively.
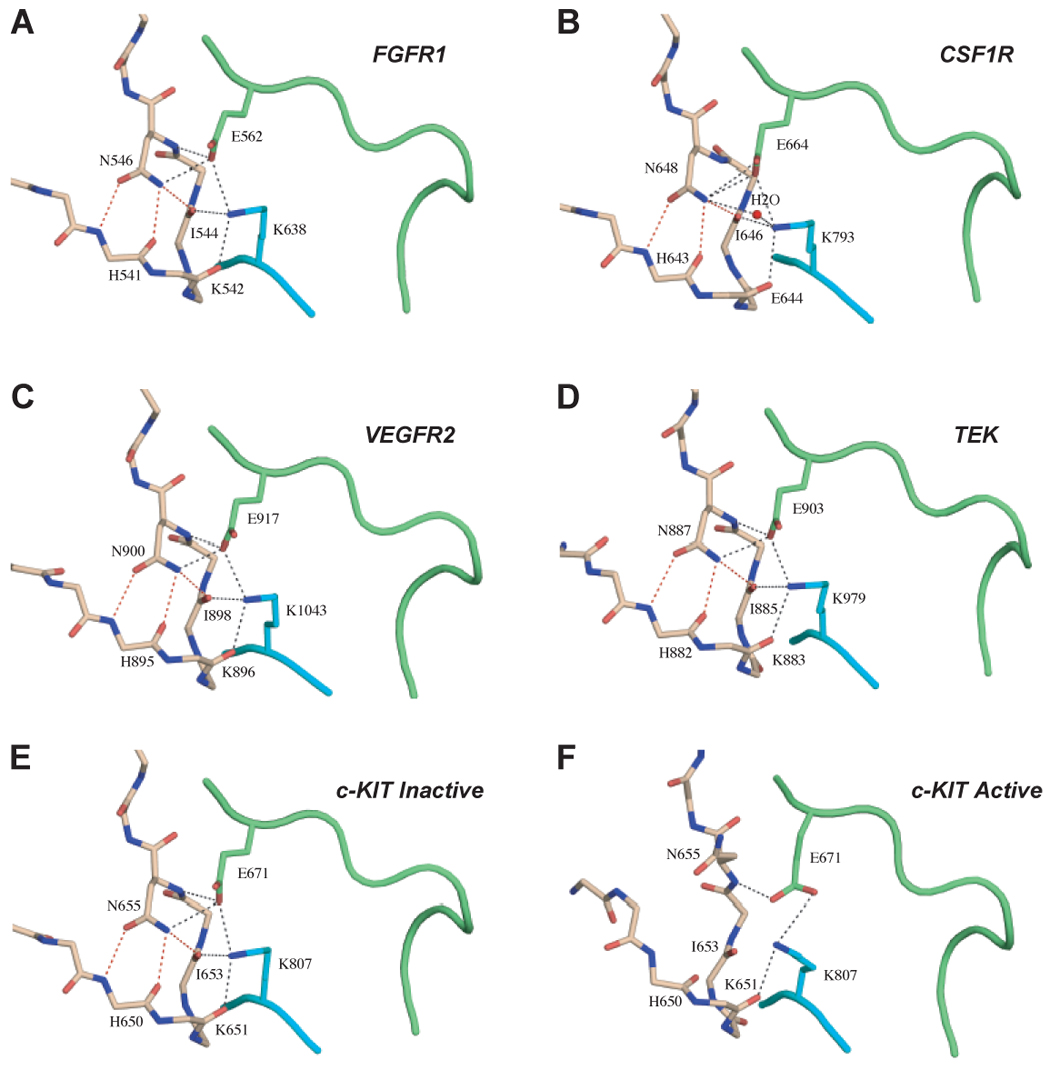
The autoinhibition by the molecular brake is a common regulatory mechanism for many RTKs. (A) to (E) show the presence of the engaged molecular brake at the kinase hinge region of unphosphorylated wild-type FGFR1 (PDB ID: 1FGK), CSF1R (PDB ID: 2I1M), VEGFR2 (PDB ID: 1VR2), TEK (PDB ID: 1FVR) and c-KIT (PDB ID: 1T45) kinases, respectively. (F) shows the disengagement of the molecular brake at the kinase hinge region of an “active” c-KIT kinase (PDB ID 1PKG). Coloring scheme is as in Fig. 3.
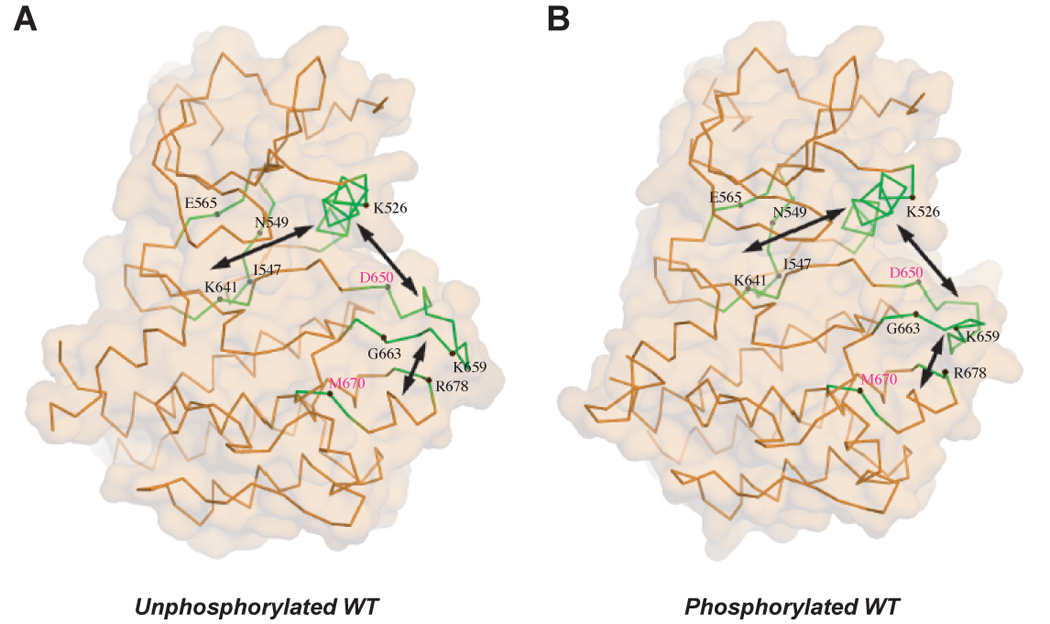
The pathogenic FGFR2K mutations highlight the existence of an allosteric communication between the three key regulatory regions of RTKs. The locations of mutated residues (labeled in black) are mapped onto the crystal structures of unphosphorylated (A) and phosphorylated (B) wild-type FGFR2 kinase. D650V and M670T mutations (residue location labeled in purple) also activate FGFR2K (unpublished results). D650V corresponds to the recurrent activating mutations D842V in PDGFRα, D816V in KIT and D835V in FLT3. M670T corresponds to the activating mutation of M918T in RET. The double headed arrows denote the reciprocal direction of communication between the A-loop, the αC helix and the kinase hinge.
References
-
- Barker SC, Kassel DB, Weigl D, Huang X, Luther MA, Knight WB. Characterization of pp60c-src tyrosine kinase activities using a continuous assay: autoactivation of the enzyme is an intermolecular autophosphorylation process. Biochemistry. 1995;34:14843–14851. - PubMed
-
- Bellus GA, McIntosh I, Smith EA, Aylsworth AS, Kaitila I, Horton WA, Greenhaw GA, Hecht JT, Francomano CA. A recurrent mutation in the tyrosine kinase domain of fibroblast growth factor receptor 3 causes hypochondroplasia. Nat Genet. 1995;10:357–359. - PubMed
-
- Bellus GA, Spector EB, Speiser PW, Weaver CA, Garber AT, Bryke CR, Israel J, Rosengren SS, Webster MK, Donoghue DJ, Francomano CA. Distinct missense mutations of the FGFR3 lys650 codon modulate receptor kinase activation and the severity of the skeletal dysplasia phenotype. Am J Hum Genet. 2000;67:1411–1421. - PMC - PubMed
-
- Brunger AT, Adams PD, Clore GM, DeLano WL, Gros P, Grosse-Kunstleve RW, Jiang JS, Kuszewski J, Nilges M, Pannu NS, et al. Crystallography & NMR system: A new software suite for macromolecular structure determination. Acta Crystallogr D Biol Crystallogr. 1998;54:905–921. - PubMed
-
- Calvert JT, Riney TJ, Kontos CD, Cha EH, Prieto VG, Shea CR, Berg JN, Nevin NC, Simpson SA, Pasyk KA, et al. Allelic and locus heterogeneity in inherited venous malformations. Hum Mol Genet. 1999;8:1279–1289. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
