

Spinal and supraspinal postural networks
- PMID: 17822773
- PMCID: PMC2204048
- DOI: 10.1016/j.brainresrev.2007.06.017
Spinal and supraspinal postural networks
Abstract
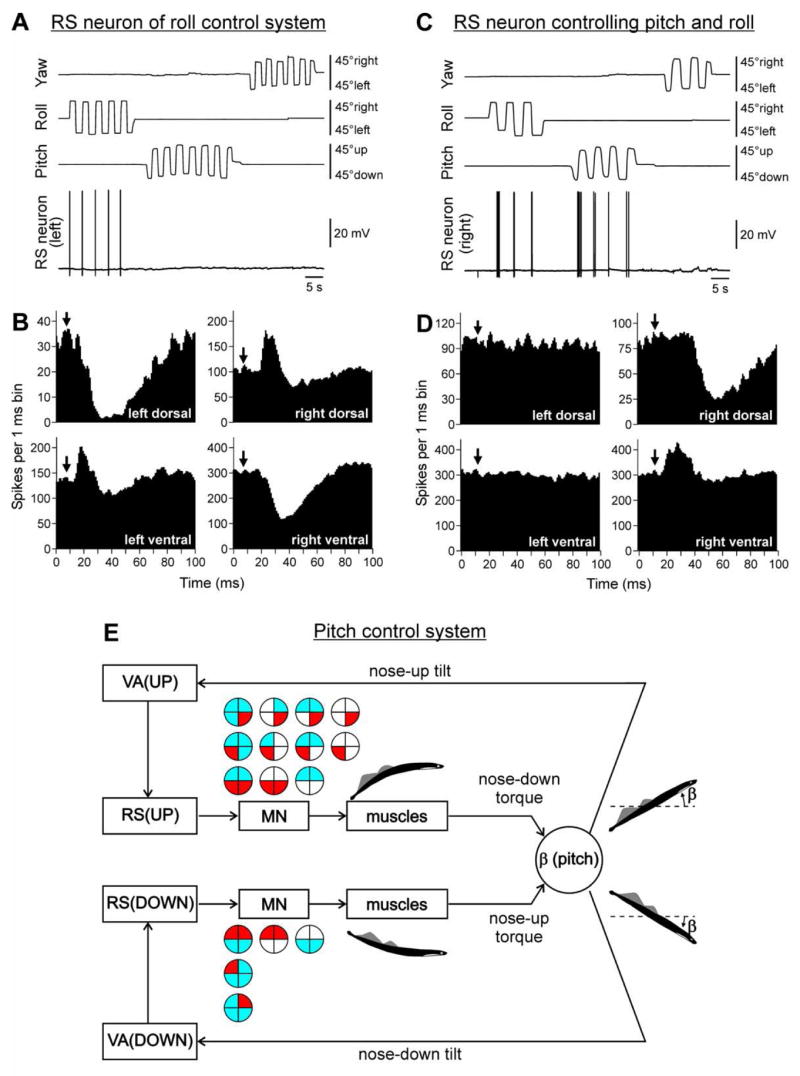
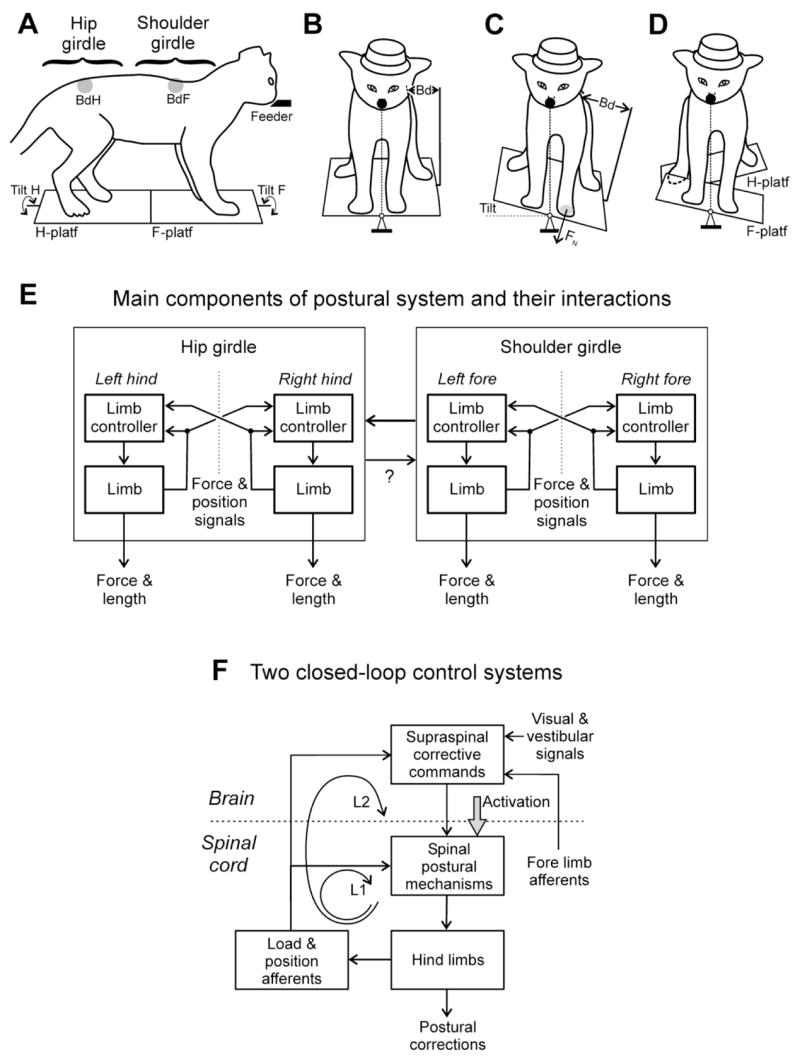
Different species maintain a particular body orientation in space (upright in humans, dorsal-side-up in quadrupeds, fish and lamprey) due to the activity of a closed-loop postural control system. We will discuss operation of spinal and supraspinal postural networks studied in a lower vertebrate (lamprey) and in two mammals (rabbit and cat). In the lamprey, the postural control system is driven by vestibular input. The key role in the postural network belongs to the reticulospinal (RS) neurons. Due to vestibular input, deviation from the stabilized body orientation in any (roll, pitch, yaw) plane leads to generation of RS commands, which are sent to the spinal cord and cause postural correction. For each of the planes, there are two groups of RS neurons responding to rotation in the opposite directions; they cause a turn opposite to the initial one. The command transmitted by an individual RS neuron causes the motor response, which contributes to the correction of posture. In each plane, the postural system stabilizes the orientation at which the antagonistic vestibular reflexes compensate for each other. Thus, in lamprey the supraspinal networks play a crucial role in stabilization of body orientation, and the function of the spinal networks is transformation of supraspinal commands into the motor pattern of postural corrections. In terrestrial quadrupeds, the postural system stabilizing the trunk orientation in the transversal plane was analyzed. It consists of two relatively independent sub-systems stabilizing orientation of the anterior and posterior parts of the trunk. They are driven by somatosensory input from limb mechanoreceptors. Each sub-system consists of two closed-loop mechanisms - spinal and spino-supraspinal. Operation of the supraspinal networks was studied by recording the posture-related activity of corticospinal neurons. The postural capacity of spinal networks was evaluated in animals with lesions to the spinal cord. Relative contribution of spinal and supraspinal mechanisms to the stabilization of trunk orientation is discussed.
Figures


References
-
- Barberini CL, Macpherson JM. Effect of head position on postural orientation and equilibrium. Exp Brain Res. 1998;122:175–184. - PubMed
-
- Bard P, Macht MB. The behavior of chronically decerebrate cat. In: Wolstenholme GEW, O’Connor CM, editors. Neurological basis of behavior. Churchill; London: 1958. pp. 55–71.
-
- Beloozerova IN, Zelenin PV, Popova LB, Orlovsky GN, Grillner S, Deliagina TG. Postural control in the rabbit maintaining balance on the tilting platform. J Neurophysiol. 2003;90:3783–3793. - PubMed
-
- Beloozerova IN, Sirota MG, Orlovsky GN, Deliagina TG. Activity of pyramidal tract neurons in the cat during postural corrections. J Neurophysiol. 2005;93:1831–1844. - PubMed
-
- Berthoz A, Pozzo T. Intermittent head stabilization during postural and locomotory tasks in humans. In: Amblard B, Berthoz A, Clarac F, editors. Posture and Gait: Development, Adaptation and Modulation. Exerpta Medica; Amsterdam: 1988. pp. 189–198.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
