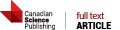Biophysical basis for airway hyperresponsiveness
- PMID: 17823634
- PMCID: PMC3901798
- DOI: 10.1139/Y07-059
Biophysical basis for airway hyperresponsiveness
Abstract
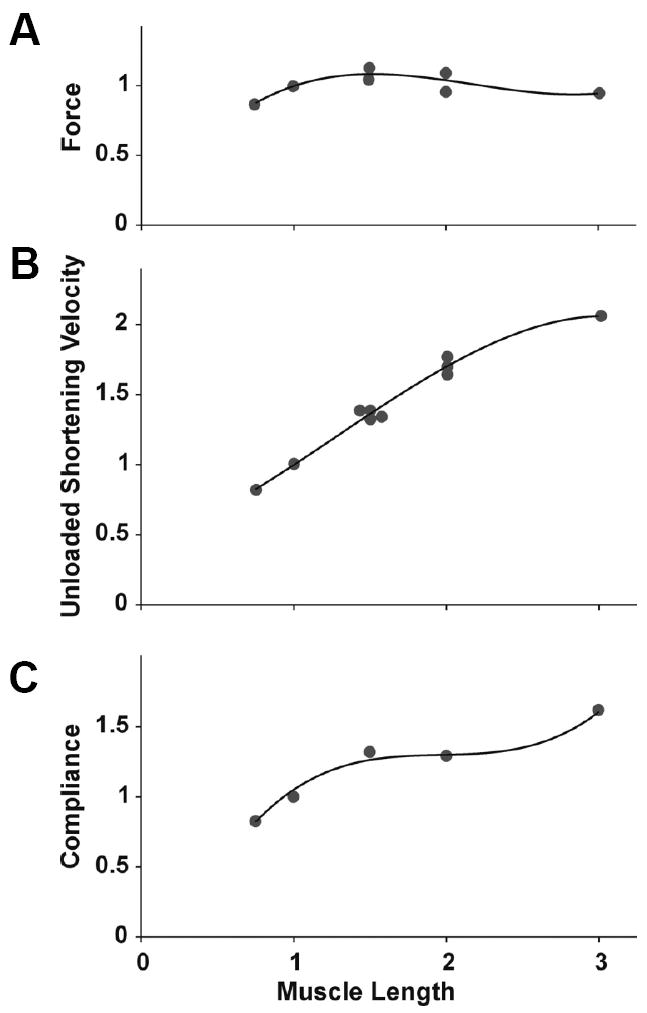
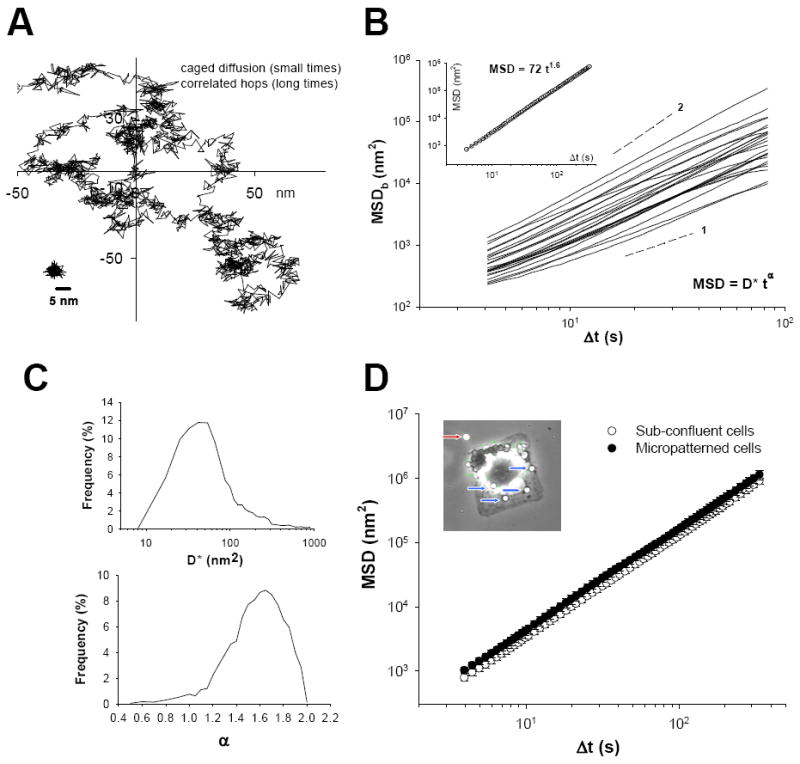
Airway hyperresponsiveness is the excessive narrowing of the airway lumen caused by stimuli that would cause little or no narrowing in the normal individual. It is one of the cardinal features of asthma, but its mechanisms remain unexplained. In asthma, the key end-effector of acute airway narrowing is contraction of the airway smooth muscle cell that is driven by myosin motors exerting their mechanical effects within an integrated cytoskeletal scaffolding. In just the past few years, however, our understanding of the rules that govern muscle biophysics has dramatically changed, as has their classical relationship to airway mechanics. It has become well established, for example, that muscle length is equilibrated dynamically rather than statically, and that in a dynamic setting nonclassical features of muscle biophysics come to the forefront, including unanticipated interactions between the muscle and its time-varying load, as well as the ability of the muscle cell to adapt (remodel) its internal microstructure rapidly in response to its ever-changing mechanical environment. Here, we consider some of these emerging concepts and, in particular, focus on structural remodeling of the airway smooth muscle cell as it relates to excessive airway narrowing in asthma.
Figures
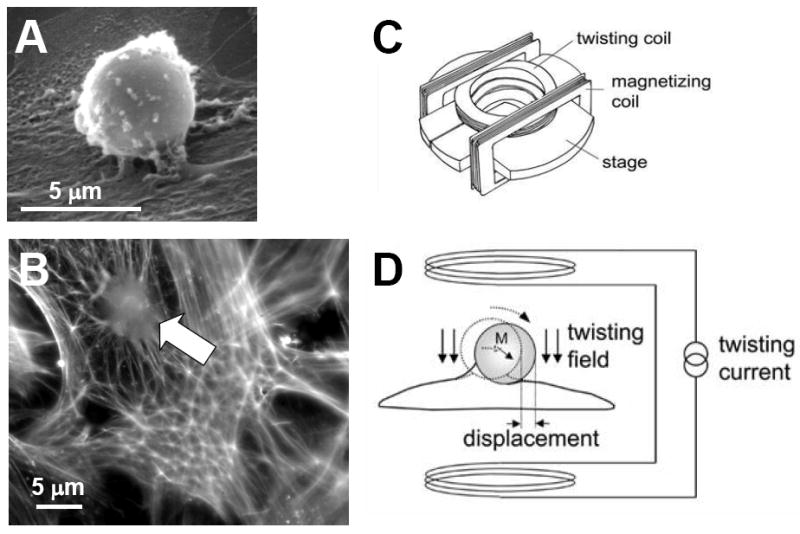
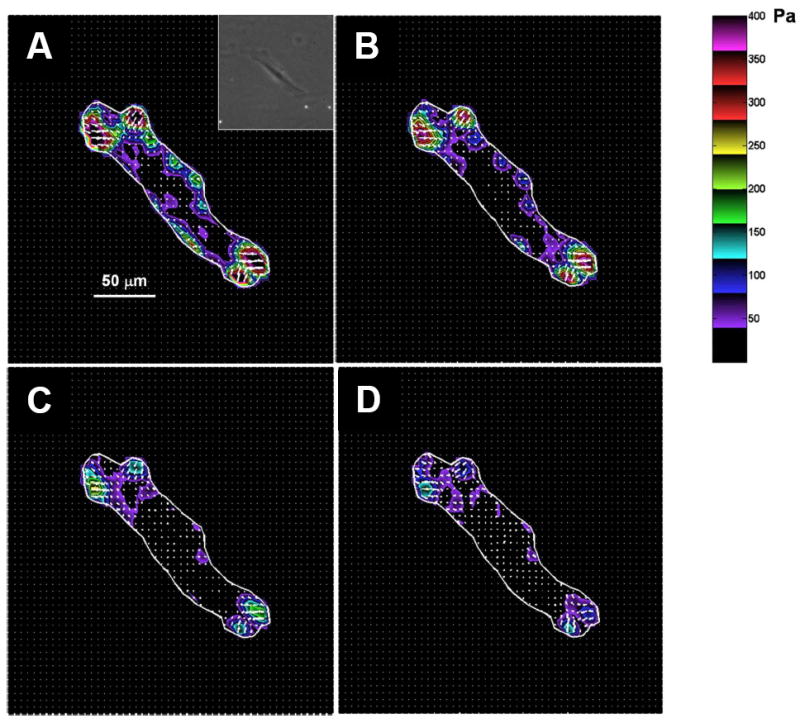
References
-
- Amrani Y, Panettieri RA. Airway smooth muscle: contraction and beyond. Internatl J Biochem Cell Biol. 2003;35:272–276. - PubMed
-
- An SS, Hai CM. Mechanical strain modulates maximal phosphatidylinositol turnover in airway smooth muscle. Am J Physiol. 1999;277:L968–L974. - PubMed
-
- An SS, Hai CM. Mechanical signals and mechanosensitive modulation of intracellular [Ca2+] in smooth muscle. Am J Physiol. 2000;279:C1375–C1384. - PubMed
-
- An SS, Laudadio RE, Lai J, Rogers RA, Fredberg JJ. Stiffness changes in cultured airway smooth muscle cells. Am J Physiol. 2002;283:C792–C801. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous