

The CCN family member Wisp3, mutant in progressive pseudorheumatoid dysplasia, modulates BMP and Wnt signaling
- PMID: 17823661
- PMCID: PMC1964511
- DOI: 10.1172/JCI32001
The CCN family member Wisp3, mutant in progressive pseudorheumatoid dysplasia, modulates BMP and Wnt signaling
Abstract
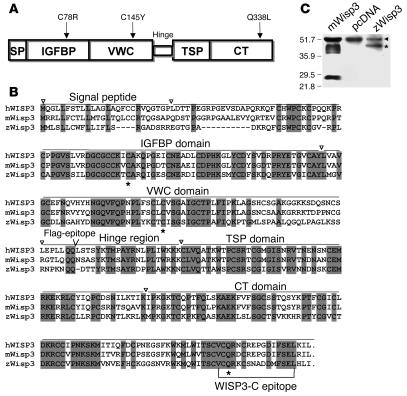
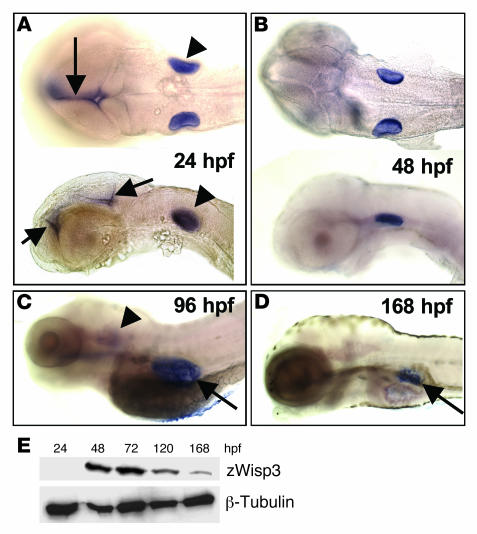
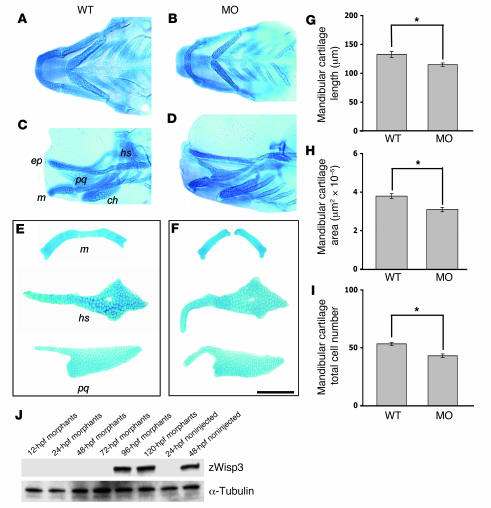
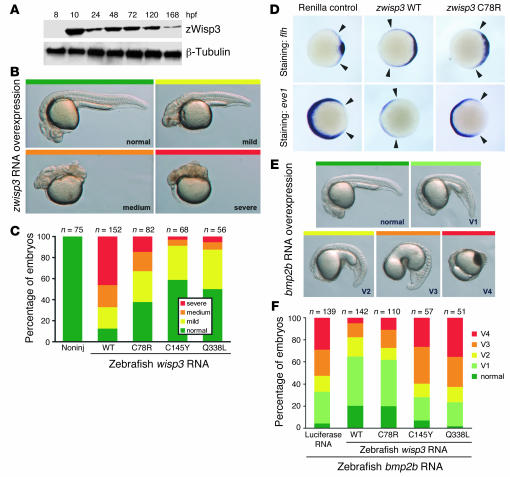
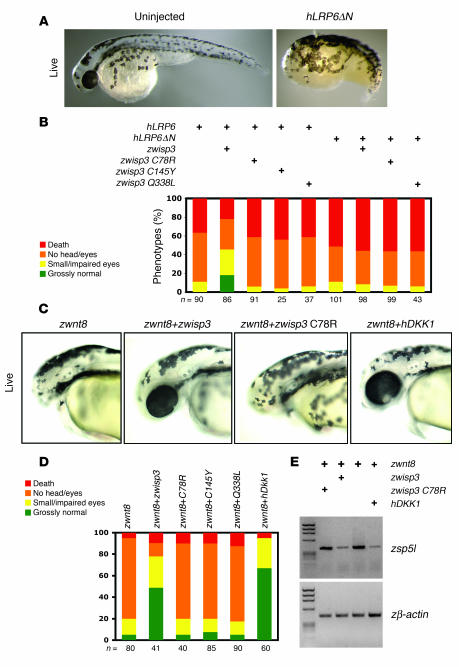
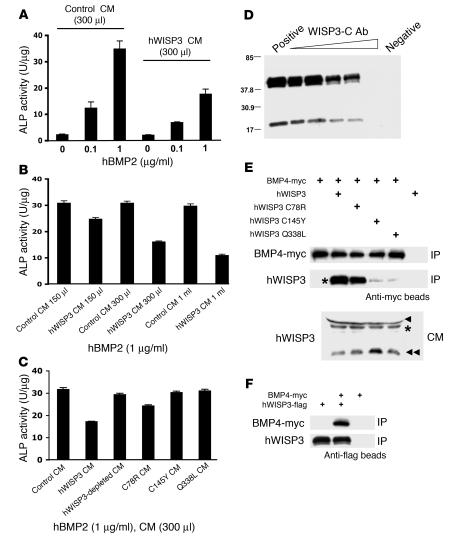
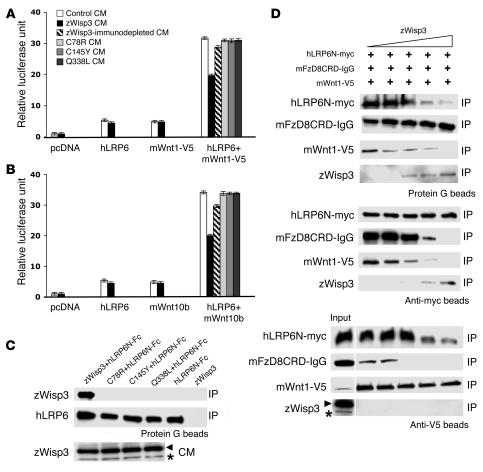
In humans, loss-of-function mutations in the gene encoding Wnt1 inducible signaling pathway protein 3 (WISP3) cause the autosomal-recessive skeletal disorder progressive pseudorheumatoid dysplasia (PPD). However, in mice there is no apparent phenotype caused by Wisp3 deficiency or overexpression. Consequently, the in vivo activities of Wisp3 have remained elusive. We cloned the zebrafish ortholog of Wisp3 and investigated its biologic activity in vivo using gain-of-function and loss-of-function approaches. Overexpression of zebrafish Wisp3 protein inhibited bone morphogenetic protein (BMP) and Wnt signaling in developing zebrafish. Conditioned medium-containing zebrafish and human Wisp3 also inhibited BMP and Wnt signaling in mammalian cells by binding to BMP ligand and to the Wnt coreceptors low-density lipoprotein receptor-related protein 6 (LRP6) and Frizzled, respectively. Wisp3 proteins containing disease-causing amino acid substitutions found in patients with PPD had reduced activity in these assays. Morpholino-mediated inhibition of zebrafish Wisp3 protein expression in developing zebrafish affected pharyngeal cartilage size and shape. These data provide a biologic assay for Wisp3, reveal a role for Wisp3 during zebrafish cartilage development, and suggest that dysregulation of BMP and/or Wnt signaling contributes to cartilage failure in humans with PPD.
Figures
Similar articles
-
WISP3, the gene responsible for the human skeletal disease progressive pseudorheumatoid dysplasia, is not essential for skeletal function in mice.Mol Cell Biol. 2005 Jan;25(1):414-21. doi: 10.1128/MCB.25.1.414-421.2005. Mol Cell Biol. 2005. PMID: 15601861 Free PMC article.
-
Mutant WISP3 triggers the phenotype shift of articular chondrocytes by promoting sensitivity to IGF-1 hypothesis of spondyloepiphyseal dysplasia tarda with progressive arthropathy (SEDT-PA).Med Hypotheses. 2007;68(6):1406-10. doi: 10.1016/j.mehy.2006.06.046. Epub 2007 Mar 23. Med Hypotheses. 2007. PMID: 17363178
-
Cellular and molecular responses in progressive pseudorheumatoid dysplasia articular cartilage associated with compound heterozygous WISP3 gene mutation.J Mol Med (Berl). 2007 Sep;85(9):985-96. doi: 10.1007/s00109-007-0193-2. Epub 2007 May 5. J Mol Med (Berl). 2007. PMID: 17483925
-
The diagnostic challenge of progressive pseudorheumatoid dysplasia (PPRD): a review of clinical features, radiographic features, and WISP3 mutations in 63 affected individuals.Am J Med Genet C Semin Med Genet. 2012 Aug 15;160C(3):217-29. doi: 10.1002/ajmg.c.31333. Epub 2012 Jul 12. Am J Med Genet C Semin Med Genet. 2012. PMID: 22791401 Review.
-
Progressive pseudorheumatoid dysplasia: a rare childhood disease.Rheumatol Int. 2019 Mar;39(3):441-452. doi: 10.1007/s00296-018-4170-6. Epub 2018 Oct 16. Rheumatol Int. 2019. PMID: 30327864 Review.
Cited by
-
Sex Differences in the Immune System Become Evident in the Perinatal Period in the Four Core Genotypes Mouse.Front Endocrinol (Lausanne). 2021 May 27;12:582614. doi: 10.3389/fendo.2021.582614. eCollection 2021. Front Endocrinol (Lausanne). 2021. PMID: 34122327 Free PMC article.
-
Dual roles of cellular communication network factor 6 (CCN6) in the invasion and metastasis of oral cancer cells to bone via binding to BMP2 and RANKL.Carcinogenesis. 2023 Dec 2;44(8-9):695-707. doi: 10.1093/carcin/bgad057. Carcinogenesis. 2023. PMID: 37590989 Free PMC article.
-
A Novel Homozygous Frameshift Mutation in CCN6 Causing Progressive Pseudorheumatoid Dysplasia (PPRD) in a Consanguineous Yemeni Family.Front Pediatr. 2019 Jun 25;7:245. doi: 10.3389/fped.2019.00245. eCollection 2019. Front Pediatr. 2019. PMID: 31294002 Free PMC article.
-
Temporal and spatial expression of CCN genes in zebrafish.Dev Dyn. 2010 Jun;239(6):1755-67. doi: 10.1002/dvdy.22279. Dev Dyn. 2010. PMID: 20503371 Free PMC article.
-
A homozygous deletion of exon 1 in WISP3 causes progressive pseudorheumatoid dysplasia in two siblings.Hum Genome Var. 2015 Dec 3;2:15049. doi: 10.1038/hgv.2015.49. eCollection 2015. Hum Genome Var. 2015. PMID: 27081554 Free PMC article.
References
-
- Hurvitz J.R., et al. Mutations in the CCN gene family member WISP3 cause progressive pseudorheumatoid dysplasia. Nat. Genet. 1999;23:94–98. - PubMed
-
- Spranger J., Albert C., Schilling F., Bartsocas C. Progressive pseudorheumatoid arthropathy of childhood (PPAC): a hereditary disorder simulating juvenile rheumatoid arthritis. Am. J. Med. Genet. 1983;14:399–401. - PubMed
-
- Wynne-Davies R., Hall C., Ansell B.M. Spondylo-epiphysial dysplasia tarda with progressive arthropathy. A “new” disorder of autosomal recessive inheritance. J. Bone Joint Surg. Br. 1982;64:442–445. - PubMed
-
- Ehl S., et al. Clinical, radiographic, and genetic diagnosis of progressive pseudorheumatoid dysplasia in a patient with severe polyarthropathy. Rheumatol. Int. 2004;24:53–56. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
