

The subtypes of nicotinic acetylcholine receptors on dopaminergic terminals of mouse striatum
- PMID: 17825262
- PMCID: PMC2735219
- DOI: 10.1016/j.bcp.2007.07.032
The subtypes of nicotinic acetylcholine receptors on dopaminergic terminals of mouse striatum
Abstract
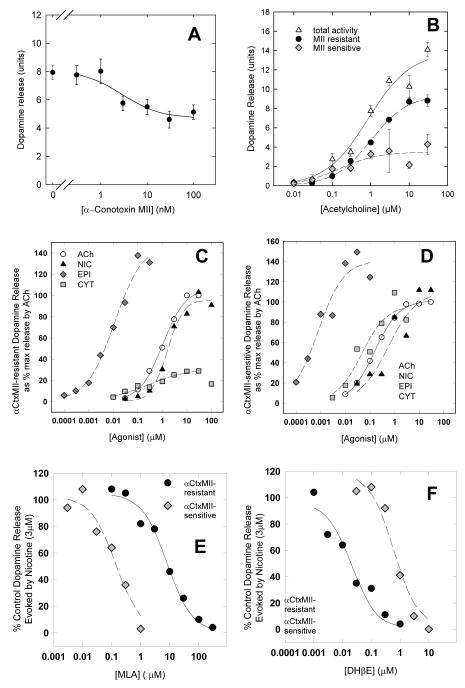
This review summarizes studies that attempted to determine the subtypes of nicotinic acetylcholine receptors (nAChR) expressed in the dopaminergic nerve terminals in the mouse. A variety of experimental approaches has been necessary to reach current knowledge of these subtypes, including in situ hybridization, agonist and antagonist binding, function measured by neurotransmitter release from synaptosomal preparations, and immunoprecipitation by selective antibodies. Early developments that facilitated this effort include the radioactive labeling of selective binding agents, such as [(125)I]-alpha-bungarotoxin and [(3)H]-nicotine, advances in cloning the subunits, and expression and evaluation of function of combinations of subunits in Xenopus oocytes. The discovery of epibatidine and alpha-conotoxin MII (alpha-CtxMII), and the development of nAChR subunit null mutant mice have been invaluable in determining which nAChR subunits are important for expression and function in mice, as well as allowing validation of the specificity of subunit specific antibodies. These approaches have identified five nAChR subtypes of nAChR that are expressed on dopaminergic nerve terminals. Three of these contain the alpha6 subunit (alpha4alpha6beta2beta3, alpha6beta2beta3, alpha6beta2) and bind alpha-CtxMII with high affinity. One of these three subtypes (alpha4alpha6beta2beta3) also has the highest sensitivity to nicotine of any native nAChR that has been studied, to date. The two subtypes that do not have high affinity for alpha-CtxMII (alpha4beta2, alpha4alpha5beta2) are somewhat more numerous than the alpha6* subtypes, but do bind nicotine with high affinity. Given that our first studies detected readily measured differences in sensitivity to agonists and antagonists among these five nAChR subtypes, it seems likely that subtype selective compounds could be developed that would allow therapeutic manipulation of diverse nAChRs that have been implicated in a number of human conditions.
Figures




References
-
- Martin LF, Kem WR, Freedman R. Alpha-7 nicotinic receptor agonists: potential new candidates for the treatment of schizophrenia. Psychopharmacology. 2004;174:54–64. - PubMed
-
- Singh A, Potter A, Newhouse P. Nicotinic acetylcholine receptor system and neuropsychiatric disorders. Idrugs. 2004;7:1096–103. - PubMed
-
- Potter AS, Newhouse PA, Bucci DJ. Central nicotinic cholinergic systems: a role in the cognitive dysfunction in attention-deficit/hyperactivity disorder? Behav Brain Research. 2006;175:201–11. - PubMed
-
- Steinlein OK. Nicotinic receptor mutations in human epilepsy. Prog in Brain Res. 2004;145:275–85. - PubMed
-
- Quik M, O’Neill M, Perez XA. Nicotine neuroprotection against nigrostriatal damage: importance of the animal model. Trends in Pharmacol Sci. 2007;28:229–35. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
