

Normal heart rhythm is initiated and regulated by an intracellular calcium clock within pacemaker cells
- PMID: 17827062
- PMCID: PMC2078332
- DOI: 10.1016/j.hlc.2007.07.005
Normal heart rhythm is initiated and regulated by an intracellular calcium clock within pacemaker cells
Abstract
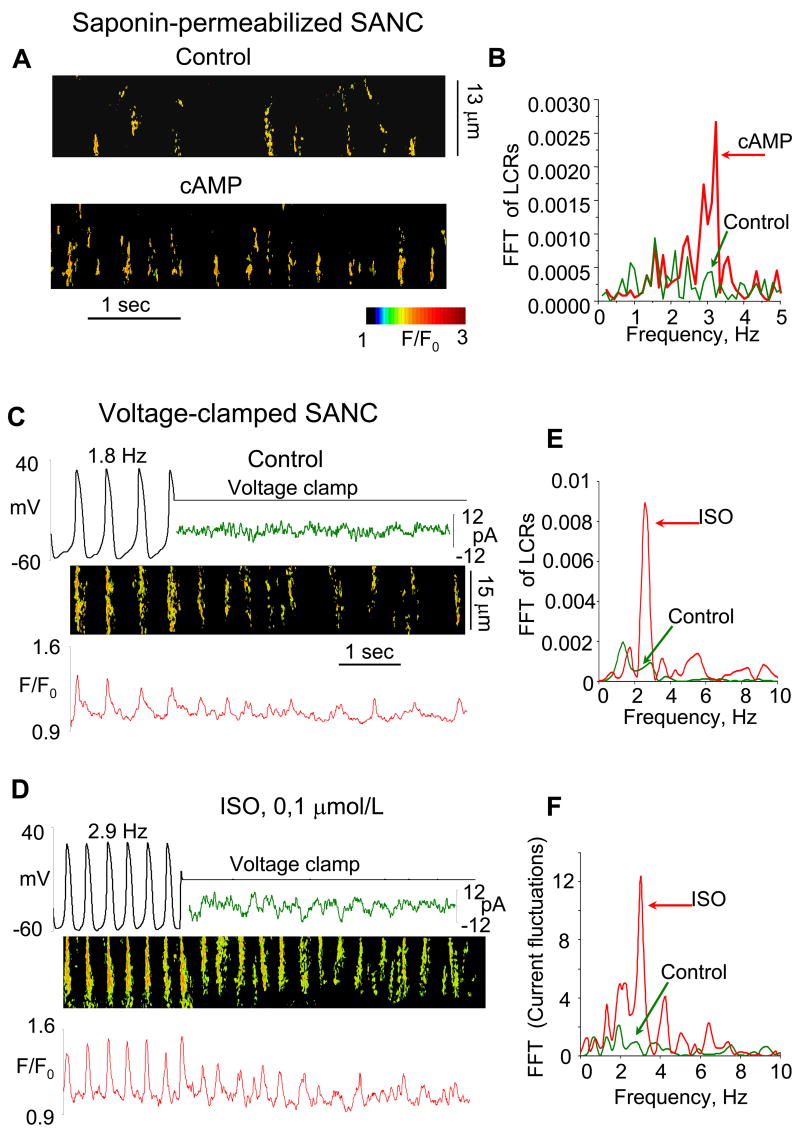
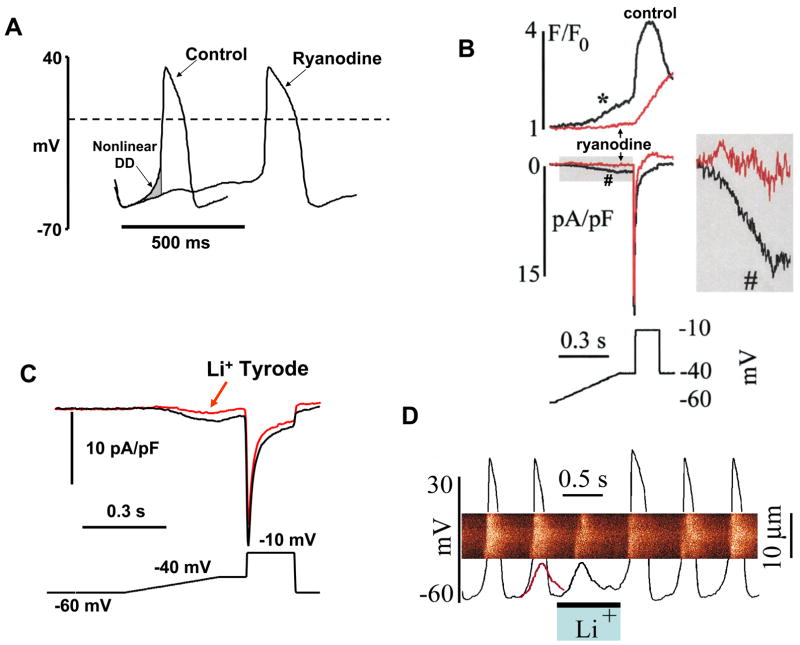
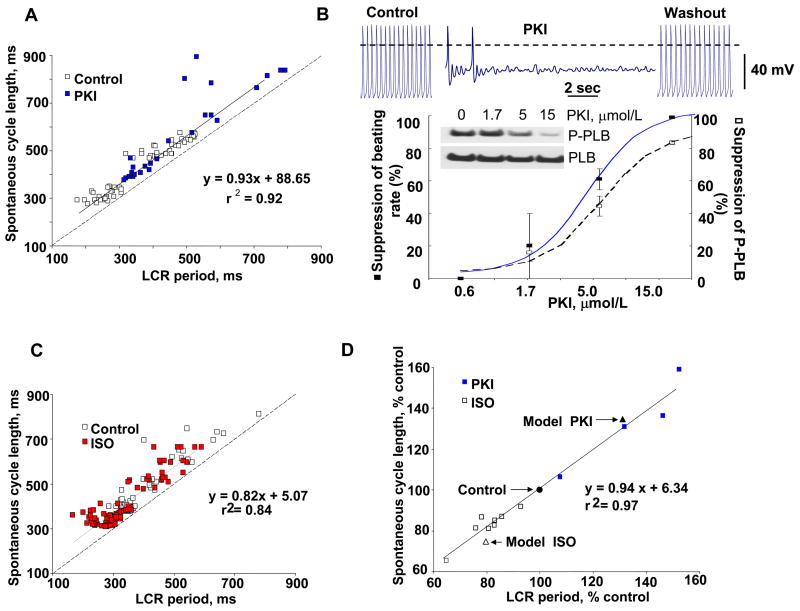
For almost half a century it has been thought that the heart rhythm originates on the surface membrane of the cardiac pacemaker cells and is driven by voltage-gated ion channels (membrane clocks). Data from several recent studies, however, conclusively show that the rhythm is initiated, sustained, and regulated by oscillatory Ca(2+) releases (Ca(2+) clock) from the sarcoplasmic reticulum, a major Ca(2+) store within sinoatrial node cells, the primary heart's pacemakers. Activation of the local oscillatory Ca(2+) releases is independent of membrane depolarisation and driven by a high level of basal state phosphorylation of Ca(2+) cycling proteins. The releases produce Ca(2+) wavelets under the cell surface membrane during the later phase of diastolic depolarisation and activate the forward mode of Na(+)/Ca(2+) exchanger resulting in inward membrane current, which ignites an action potential. Phosphorylation-dependent gradation of speed at which Ca(2+) clock cycles is the essential regulatory mechanism of normal pacemaker rate and rhythm. The robust regulation of pacemaker function is insured by tight integration of Ca(2+) and membrane clocks: the action potential shape and ion fluxes are tuned by membrane clocks to sustain operation of the Ca(2+) clock which produces timely and powerful ignition of the membrane clocks to effect action potentials.
Figures








References
-
- Cranefield PF. Action potentials, after potentials, and arrhythmias. Circ Res. 1977;41:415–23. - PubMed
-
- Maltsev VA, Vinogradova TM, Lakatta EG. The emergence of a general theory of the initiation and strength of the heartbeat. J Pharmacol Sci. 2006;100:338–69. - PubMed
-
- Noble D. Cardiac action and pacemaker potentials based on the Hodgkin-Huxley equations. Nature. 1960;188:495–7. - PubMed
-
- Vinogradova TM, Lyashkov AE, Zhu W, Ruknudin AM, Sirenko S, Yang D, Deo S, Barlow M, Johnson S, Caffrey JL, Zhou YY, Xiao RP, Cheng H, Stern MD, Maltsev VA, Lakatta EG. High basal protein kinase A-dependent phosphorylation drives rhythmic internal Ca2+ store oscillations and spontaneous beating of cardiac pacemaker cells. Circ Res. 2006;98:505–14. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
