

Cuckoos, cowbirds and hosts: adaptations, trade-offs and constraints
- PMID: 17827098
- PMCID: PMC2442387
- DOI: 10.1098/rstb.2006.1849
Cuckoos, cowbirds and hosts: adaptations, trade-offs and constraints
Abstract
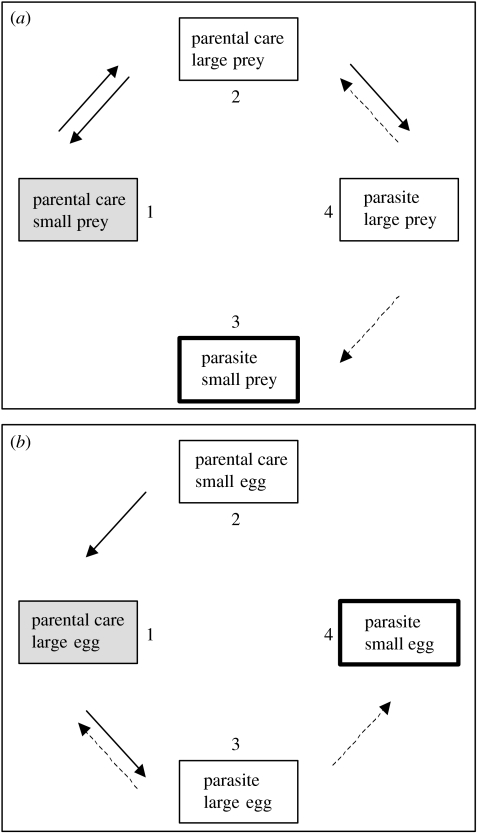
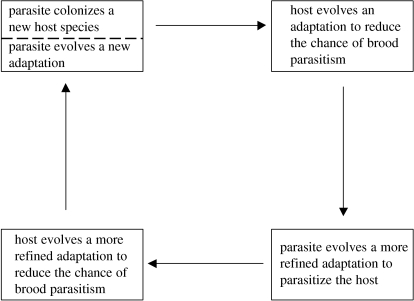
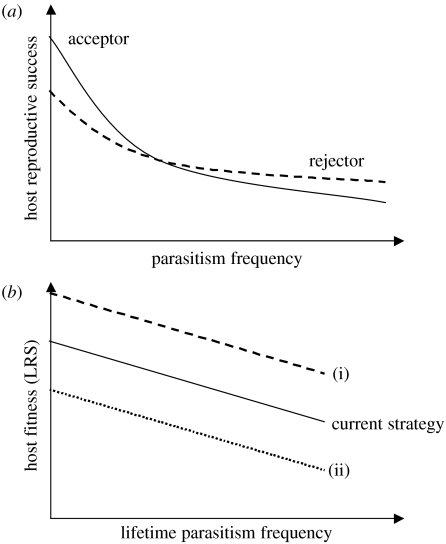
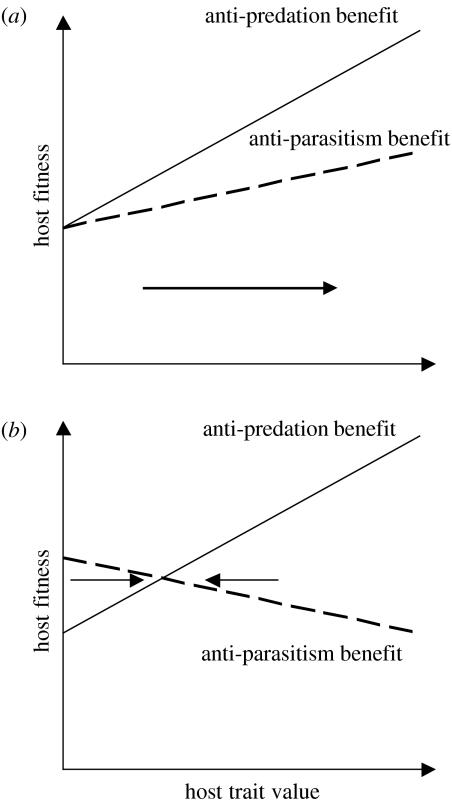
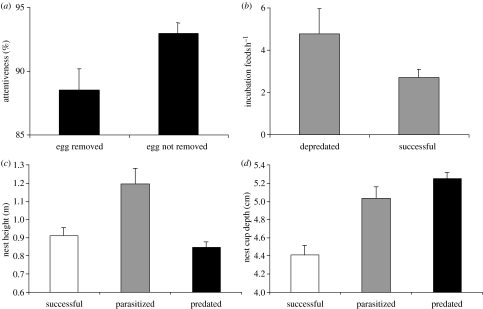
The interactions between brood parasitic birds and their host species provide one of the best model systems for coevolution. Despite being intensively studied, the parasite-host system provides ample opportunities to test new predictions from both coevolutionary theory as well as life-history theory in general. I identify four main areas that might be especially fruitful: cuckoo female gentes as alternative reproductive strategies, non-random and nonlinear risks of brood parasitism for host individuals, host parental quality and targeted brood parasitism, and differences and similarities between predation risk and parasitism risk. Rather than being a rare and intriguing system to study coevolutionary processes, I believe that avian brood parasites and their hosts are much more important as extreme cases in the evolution of life-history strategies. They provide unique examples of trade-offs and situations where constraints are either completely removed or particularly severe.
Figures
References
-
- Alderson G.W, Gibbs H.L, Sealy S.G. Determining the reproductive behaviour of individual brown-headed cowbirds using microsatellite DNA markers. Anim. Behav. 1999;58:895–905. doi:10.1006/anbe.1999.1220 - DOI - PubMed
-
- Alvarez F. Proximity of trees facilitates parasitism by cuckoos Cuculus canorus on rufous warblers Cercotrichas galactotes. Ibis. 1993;135:331.
-
- Amundsen T, Brobakken P.T, Moksnes A, Røskaft E. Rejection of common cuckoo Cuculus canorus eggs in relation to female age in the bluethroat Luscinia svecica. J. Avian Biol. 2002;33:366–370. doi:10.1034/j.1600-048X.2002.02894.x - DOI
-
- Aragon S, Møller A.P, Soler J.J, Soler M. Molecular phylogeny of cuckoos supports a polyphyletic orgin of brood parasitism. J. Evol. Biol. 1999;12:495–506. doi:10.1046/j.1420-9101.1999.00052.x - DOI
-
- Arcese P, Smith J.N.M, Hatch M.I. Nest predation by cowbirds and its consequences for passerine demography. Proc. Natl Acad. Sci. USA. 1996;93:4608–4611. doi:10.1073/pnas.93.10.4608 - DOI - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources