

Hemoglobin is a co-factor of human trypanosome lytic factor
- PMID: 17845074
- PMCID: PMC1971115
- DOI: 10.1371/journal.ppat.0030129
Hemoglobin is a co-factor of human trypanosome lytic factor
Abstract
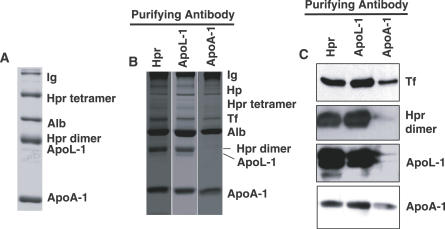
Trypanosome lytic factor (TLF) is a high-density lipoprotein (HDL) subclass providing innate protection to humans against infection by the protozoan parasite Trypanosoma brucei brucei. Two primate-specific plasma proteins, haptoglobin-related protein (Hpr) and apolipoprotein L-1 (ApoL-1), have been proposed to kill T. b. brucei both singularly or when co-assembled into the same HDL. To better understand the mechanism of T. b. brucei killing by TLF, the protein composition of TLF was investigated using a gentle immunoaffinity purification technique that avoids the loss of weakly associated proteins. HDL particles recovered by immunoaffinity absorption, with either anti-Hpr or anti-ApoL-1, were identical in protein composition and specific activity for T. b. brucei killing. Here, we show that TLF-bound Hpr strongly binds Hb and that addition of Hb stimulates TLF killing of T. b. brucei by increasing the affinity of TLF for its receptor, and by inducing Fenton chemistry within the trypanosome lysosome. These findings suggest that TLF in uninfected humans may be inactive against T. b. brucei prior to initiation of infection. We propose that infection of humans by T. b. brucei causes hemolysis that triggers the activation of TLF by the formation of Hpr-Hb complexes, leading to enhanced binding, trypanolytic activity, and clearance of parasites.
Conflict of interest statement
Figures






References
-
- Hoare CA. The Trypanosomes of mammals. A zoological monograph. Oxford: Blackwell Scientific Publications; 1972.
-
- Gibson W. Will the real Trypanosoma brucei rhodesiense please step forward? Trends Parasitol. 2002;18:486–490. - PubMed
-
- Gibson WC. Will the real Trypanosoma b. gambiense please stand up. Parasitol Today. 1986;2:255–257. - PubMed
-
- Pays E, Vanhamme L, Perez-Morga D. Antigenic variation in Trypanosoma brucei: Facts, challenges and mysteries. Curr Opin Microbiol. 2004;7:369–374. - PubMed
-
- Hajduk SL, Hager K, Esko JD. High-density lipoprotein-mediated lysis of trypanosomes. Parasitol Today. 1992;8:95–98. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
