

Suv4-20h deficiency results in telomere elongation and derepression of telomere recombination
- PMID: 17846168
- PMCID: PMC2064618
- DOI: 10.1083/jcb.200703081
Suv4-20h deficiency results in telomere elongation and derepression of telomere recombination
Abstract
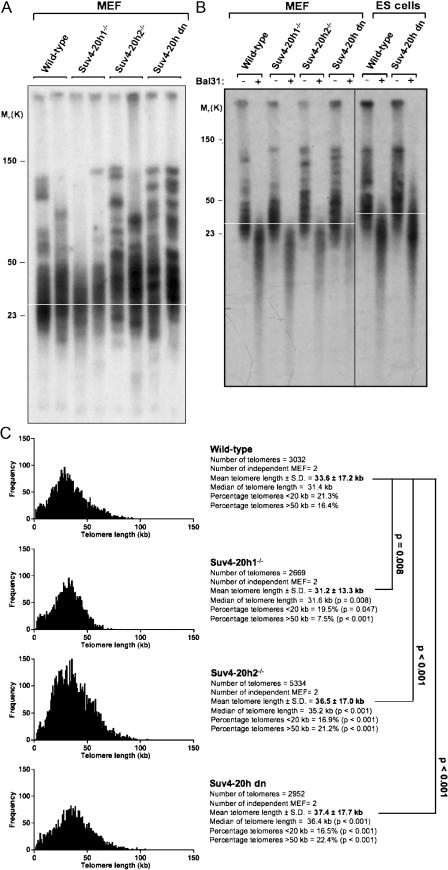
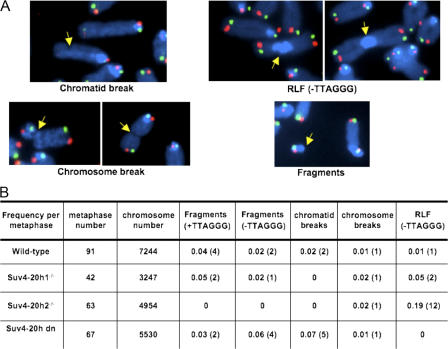
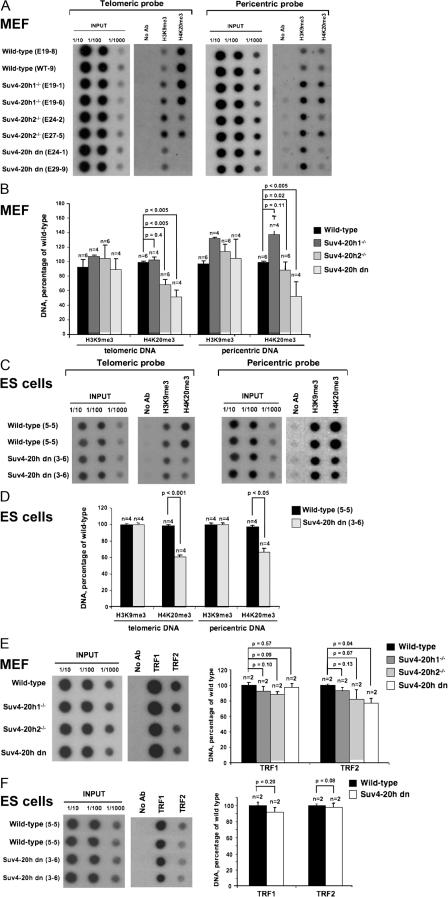
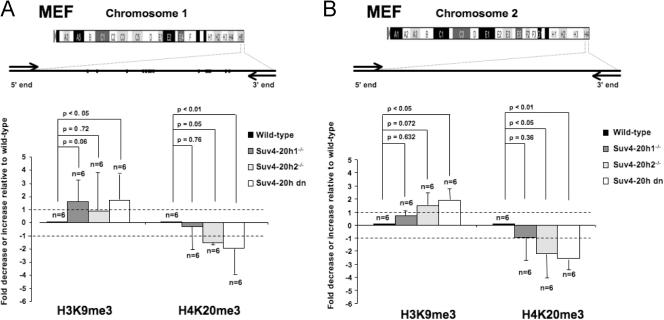
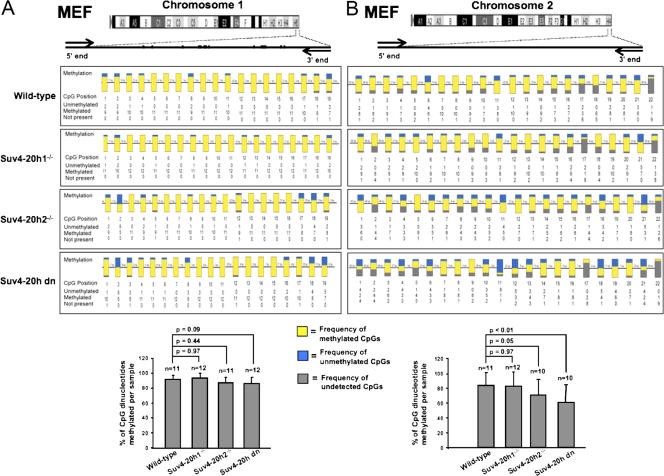
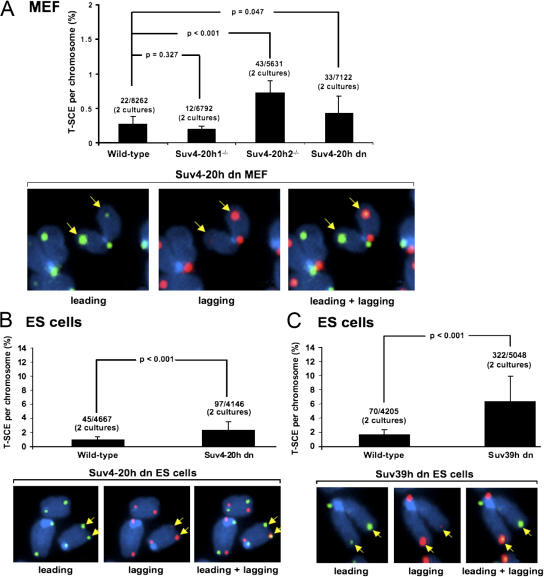
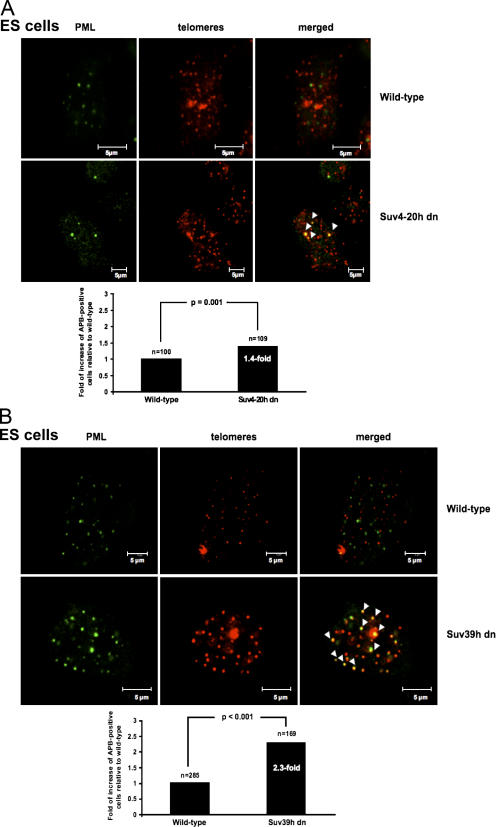
Mammalian telomeres have heterochromatic features, including trimethylated histone H3 at lysine 9 (H3K9me3) and trimethylated histone H4 at lysine 20 (H4K20me3). In addition, subtelomeric DNA is hypermethylated. The enzymatic activities responsible for these modifications at telomeres are beginning to be characterized. In particular, H4K20me3 at telomeres could be catalyzed by the novel Suv4-20h1 and Suv4-20h2 histone methyltransferases (HMTases). In this study, we demonstrate that the Suv4-20h enzymes are responsible for this histone modification at telomeres. Cells deficient for Suv4-20h2 or for both Suv4-20h1 and Suv4-20h2 show decreased levels of H4K20me3 at telomeres and subtelomeres in the absence of changes in H3K9me3. These epigenetic alterations are accompanied by telomere elongation, indicating a role for Suv4-20h HMTases in telomere length control. Finally, cells lacking either the Suv4-20h or Suv39h HMTases show increased frequencies of telomere recombination in the absence of changes in subtelomeric DNA methylation. These results demonstrate the importance of chromatin architecture in the maintenance of telomere length homeostasis and reveal a novel role for histone lysine methylation in controlling telomere recombination.
Figures
References
-
- Bailey, S.M., M.N. Cornforth, A. Kurimasa, D.J. Chen, and E.H. Goodwin. 2001. Strand-specific postreplicative processing of mammalian telomeres. Science. 293:2462–2465. - PubMed
-
- Bechter, O.E., Y. Zou, W. Walker, W.E. Wright, and J.W. Shay. 2004. Telomeric recombination in mismatch repair deficient human colon cancer cells after telomerase inhibition. Cancer Res. 64:3444–3451. - PubMed
-
- Benetti, R., M. García-Cao, and M.A. Blasco. 2007. Telomere length regulates the epigenetic status of mammalian telomeres and subtelomeres. Nat. Genet. 39:243–250. - PubMed
-
- Blasco, M.A. 2005. Telomeres and human disease: ageing, cancer and beyond. Nat. Rev. Genet. 6:611–622. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
