

Restricted active site docking by enzyme-bound substrate enforces the ordered cleavage of prothrombin by prothrombinase
- PMID: 17848548
- PMCID: PMC2292459
- DOI: 10.1074/jbc.M706529200
Restricted active site docking by enzyme-bound substrate enforces the ordered cleavage of prothrombin by prothrombinase
Abstract
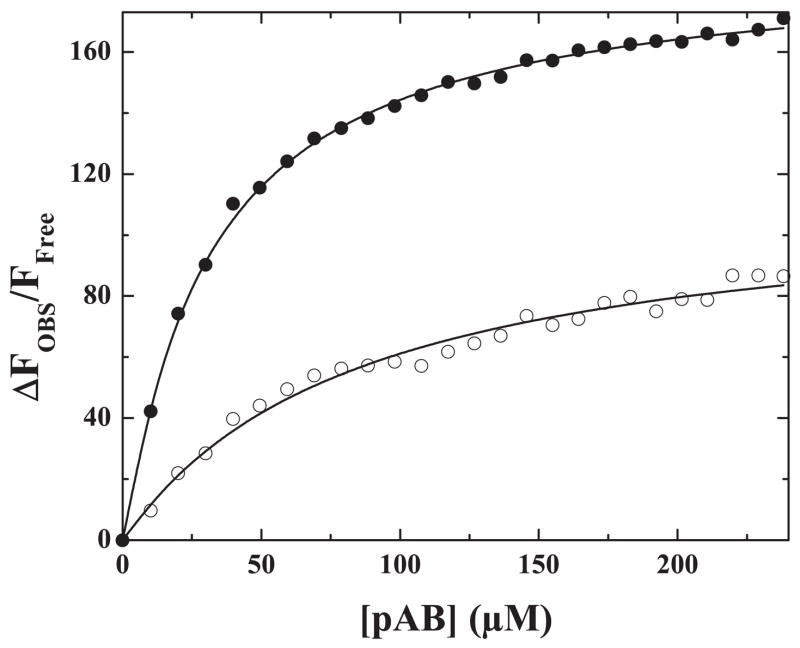
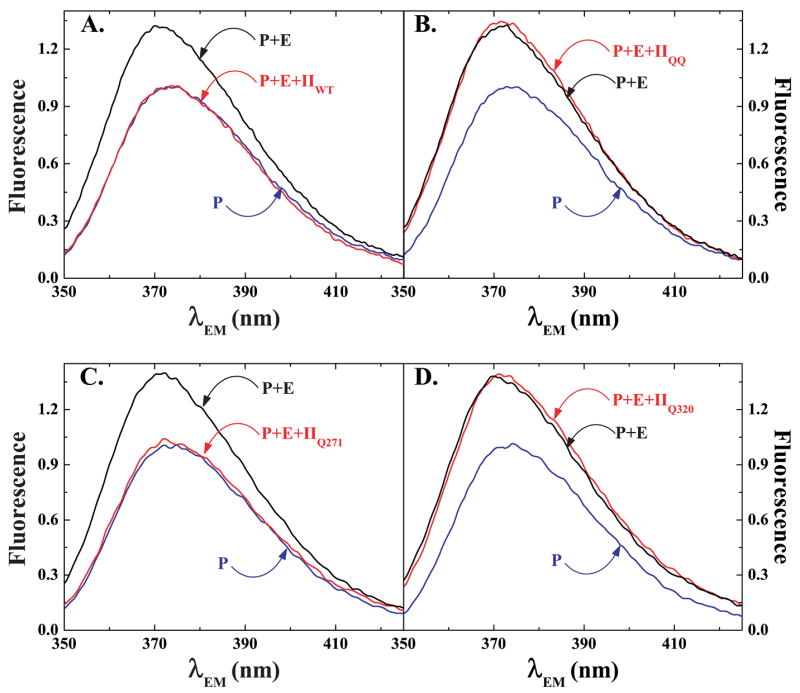
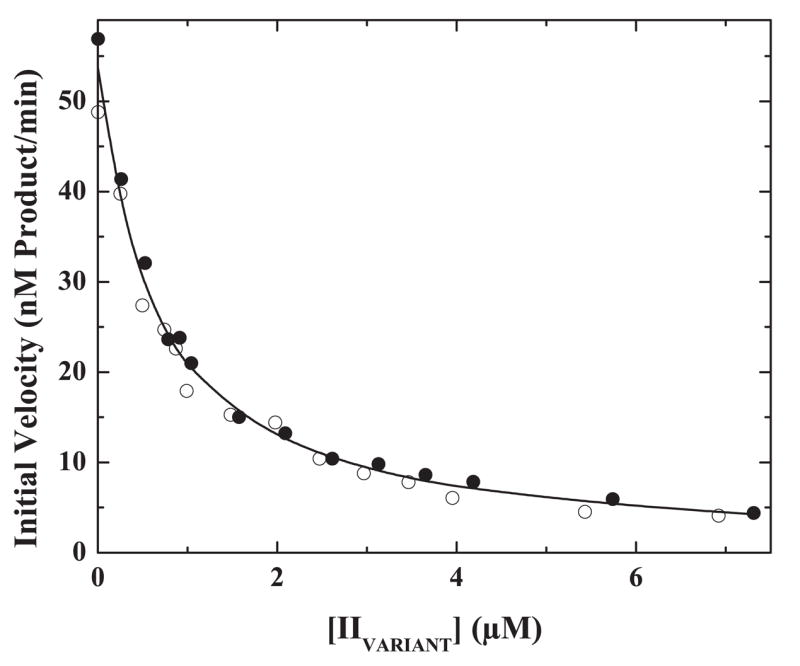
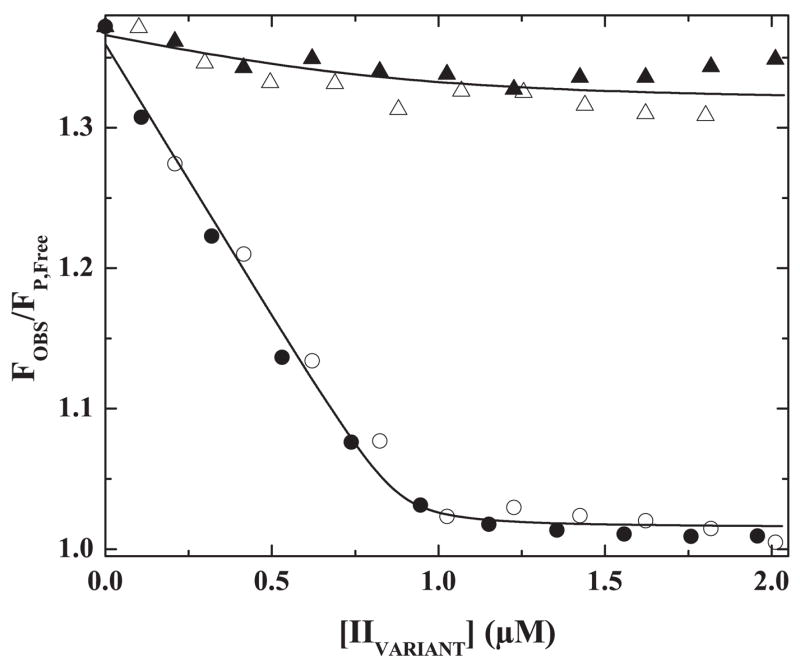
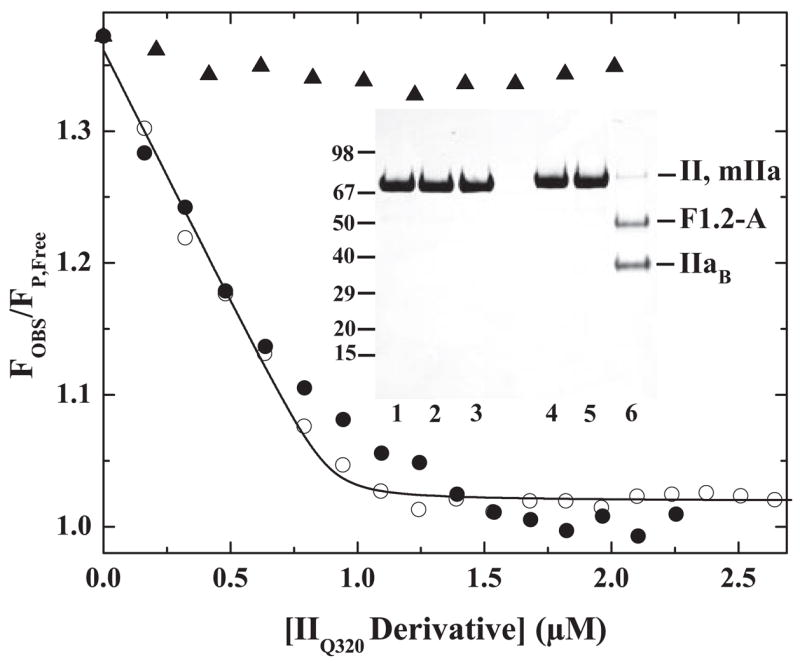
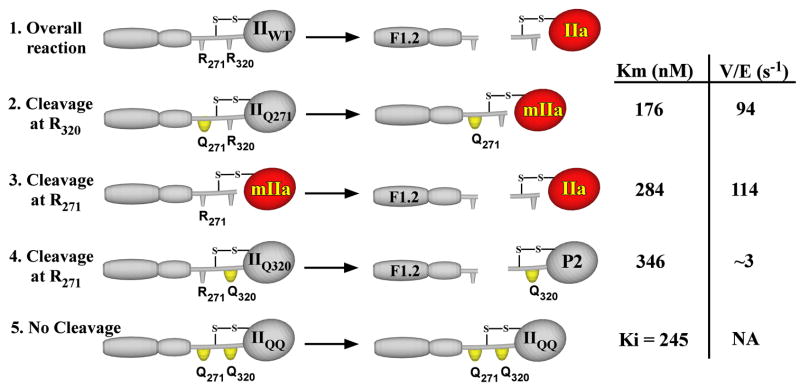
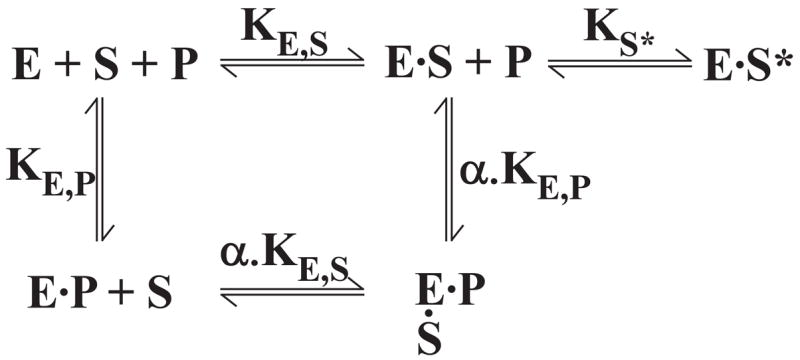
The preferred pathway for prothrombin activation by prothrombinase involves initial cleavage at Arg(320) to produce meizothrombin, which is then cleaved at Arg(271) to liberate thrombin. Exosite binding drives substrate affinity and is independent of the bond being cleaved. The pathway for cleavage is determined by large differences in V(max) for cleavage at the two sites within intact prothrombin. By fluorescence binding studies in the absence of catalysis, we have assessed the ability of the individual cleavage sites to engage the active site of Xa within prothrombinase at equilibrium. Using a panel of recombinant cleavage site mutants, we show that in intact prothrombin, the Arg(320) site effectively engages the active site in a 1:1 interaction between substrate and enzyme. In contrast, the Arg(271) site binds to the active site poorly in an interaction that is approximately 600-fold weaker. Perceived substrate affinity is independent of active site engagement by either cleavage site. We further show that prior cleavage at the 320 site or the stabilization of the uncleaved zymogen in a proteinase-like state facilitates efficient docking of Arg(271) at the active site of prothrombinase. Therefore, we establish direct relationships between docking of either cleavage site at the active site of the catalyst, the V(max) for cleavage at that site, substrate conformation, and the resulting pathway for prothrombin cleavage. Exosite tethering of the substrate in either the zymogen or proteinase conformation dictates which cleavage site can engage the active site of the catalyst and enforces the sequential cleavage of prothrombin by prothrombinase.
Figures
References
-
- Jackson CM, Nemerson Y. Annu Rev Biochem. 1980;49:765–811. - PubMed
-
- Mann KG, Jenny RJ, Krishnaswamy S. Annu Rev Biochem. 1988;57:915–956. - PubMed
-
- Mann KG, Nesheim ME, Church WR, Haley P, Krishnaswamy S. Blood. 1990;76:1–16. - PubMed
-
- Camire RM, Pollack ES. In: Hemostasis and Thrombosis, Basic Principles and Clinical Practice. Colman RW, Marder VJ, Clowes AJ, George JN, Goldhaber SZ, editors. Lippincott Williams & Wilkins; Philadelphia: 2006. pp. 59–89.
-
- Jenny NS, Lundblad RL, Mann KG. In: Hemostasis and Thrombosis, Basic Principles and Clinical Practice. Colman RW, Marder VJ, Clowes AJ, George JN, Goldhaber SZ, editors. Lippincott Williams & Wilkins; Philadelphia: 2006. pp. 193–213.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
