

The coupling of synthesis and partitioning of EBV's plasmid replicon is revealed in live cells
- PMID: 17853891
- PMCID: PMC2000340
- DOI: 10.1038/sj.emboj.7601853
The coupling of synthesis and partitioning of EBV's plasmid replicon is revealed in live cells
Abstract
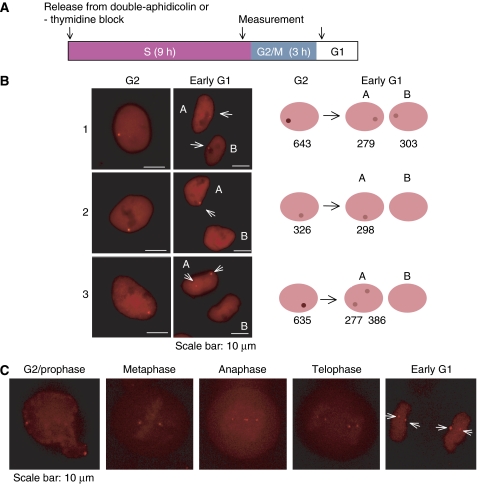
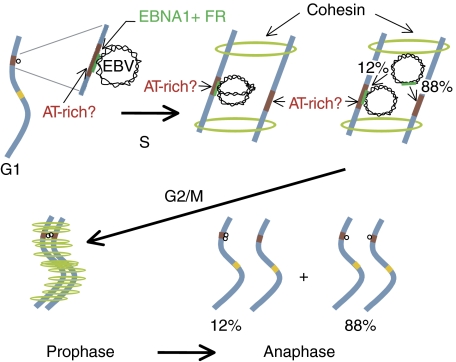
Epstein-Barr virus (EBV) is an exceptionally successful human viral pathogen maintained as a licensed, plasmid replicon in proliferating cells. We have measured the distributions of EBV-derived plasmids in single live cells throughout the cell cycle in the absence of selection and confirmed the measured rates of duplication and partitioning computationally and experimentally. These analyses have uncovered a striking, non-random partitioning for this minimalist plasmid replicon and revealed additional properties of it and its host cells: (1) 84% of the plasmids duplicate during each S phase; (2) all duplicated plasmids are spatially colocalized as pairs, a positioning that is coupled to their non-random partitioning; (3) each clone of cells requires a certain threshold number of plasmids per cell for its optimal growth under selection; (4) defects in plasmid synthesis and partitioning are balanced to yield wide distributions of plasmids in clonal populations of cells for which the plasmids provide a selective advantage. These properties of its plasmid replicon underlie EBV's success as a human pathogen.
Figures




Similar articles
-
The plasmid replicon of EBV consists of multiple cis-acting elements that facilitate DNA synthesis by the cell and a viral maintenance element.EMBO J. 1998 Nov 2;17(21):6394-403. doi: 10.1093/emboj/17.21.6394. EMBO J. 1998. PMID: 9799247 Free PMC article.
-
EBV's plasmid replicon: an enigma in cis and trans.Curr Top Microbiol Immunol. 2001;258:3-11. doi: 10.1007/978-3-642-56515-1_1. Curr Top Microbiol Immunol. 2001. PMID: 11443865 Review. No abstract available.
-
Plasmid Partitioning by Human Tumor Viruses.J Virol. 2018 Apr 13;92(9):e02170-17. doi: 10.1128/JVI.02170-17. Print 2018 May 1. J Virol. 2018. PMID: 29467315 Free PMC article. Review.
-
Replication of Epstein-Barr viral DNA.Cold Spring Harb Perspect Biol. 2013 Jan 1;5(1):a013029. doi: 10.1101/cshperspect.a013029. Cold Spring Harb Perspect Biol. 2013. PMID: 23284049 Free PMC article. Review.
-
The plasmid replicon of Epstein-Barr virus: mechanistic insights into efficient, licensed, extrachromosomal replication in human cells.Plasmid. 2007 Jul;58(1):1-12. doi: 10.1016/j.plasmid.2007.01.003. Epub 2007 Mar 9. Plasmid. 2007. PMID: 17350094 Free PMC article. Review.
Cited by
-
The role of Epstein-Barr virus infection in the pathogenesis of nasopharyngeal carcinoma.Virol Sin. 2015 Apr;30(2):107-21. doi: 10.1007/s12250-015-3592-5. Epub 2015 Apr 21. Virol Sin. 2015. PMID: 25910483 Free PMC article. Review.
-
Epstein-Barr Virus Limits the Accumulation of IPO7, an Essential Gene Product.Front Microbiol. 2021 Feb 16;12:643327. doi: 10.3389/fmicb.2021.643327. eCollection 2021. Front Microbiol. 2021. PMID: 33664726 Free PMC article.
-
Molecular and cellular characterization of an AT-hook protein from Leishmania.PLoS One. 2011;6(6):e21412. doi: 10.1371/journal.pone.0021412. Epub 2011 Jun 23. PLoS One. 2011. PMID: 21731738 Free PMC article.
-
An episomal vector-based CRISPR/Cas9 system for highly efficient gene knockout in human pluripotent stem cells.Sci Rep. 2017 May 24;7(1):2320. doi: 10.1038/s41598-017-02456-y. Sci Rep. 2017. PMID: 28539611 Free PMC article.
-
Chromatin organization of gammaherpesvirus latent genomes.Biochim Biophys Acta. 2010 Mar-Apr;1799(3-4):236-45. doi: 10.1016/j.bbagrm.2009.10.004. Epub 2009 Oct 22. Biochim Biophys Acta. 2010. PMID: 19853673 Free PMC article. Review.
References
-
- Chow LT, Broker TR (2006) Papillomavirus. In DNA Replication and Human Disease, DePamphilis ML (ed), pp 609–625. Cold Spring Harbor, New York, USA: Cold Spring Harbor Press
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
