

Highly pathogenic avian influenza H5N1 viruses elicit an attenuated type i interferon response in polarized human bronchial epithelial cells
- PMID: 17855549
- PMCID: PMC2169033
- DOI: 10.1128/JVI.01134-07
Highly pathogenic avian influenza H5N1 viruses elicit an attenuated type i interferon response in polarized human bronchial epithelial cells
Abstract
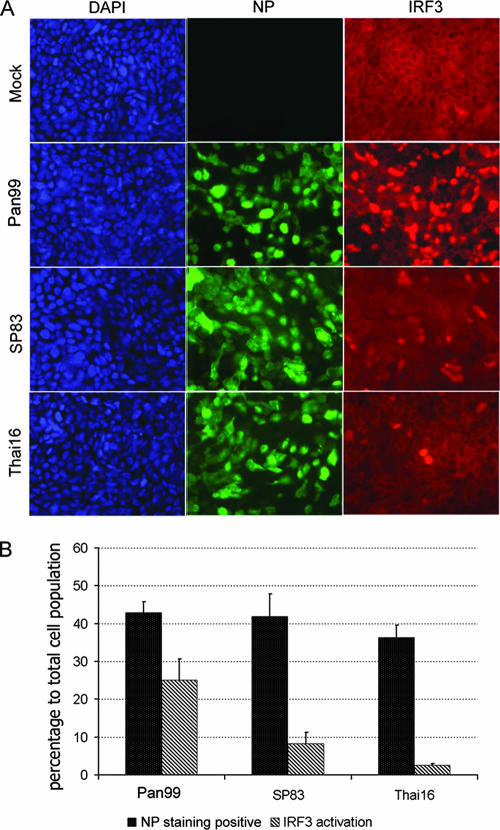
The unparalleled spread of highly pathogenic avian influenza A (HPAI) H5N1 viruses has resulted in devastating outbreaks in domestic poultry and sporadic human infections with a high fatality rate. To better understand the mechanism(s) of H5N1 virus pathogenesis and host responses in humans, we utilized a polarized human bronchial epithelial cell model that expresses both avian alpha-2,3- and human alpha-2,6-linked sialic acid receptors on the apical surface and supports productive replication of both H5N1 and H3N2 viruses. Using this model, we compared the abilities of selected 2004 HPAI H5N1 viruses isolated from humans and a recent human H3N2 virus to trigger the type I interferon (IFN) response. H5N1 viruses elicited significantly less IFN regulatory factor 3 (IRF3) nuclear translocation, as well as delayed and reduced production of IFN-beta compared with the H3N2 virus. Furthermore, phosphorylation of Stat2 and induction of IFN-stimulated genes (ISGs), such as MX1, ISG15, IRF7, and retinoic acid-inducible gene I, were substantially delayed and reduced in cells infected with H5N1 viruses. We also observed that the highly virulent H5N1 virus replicated more efficiently and induced a weaker IFN response than the H5N1 virus that exhibited low virulence in mammals in an earlier study. Our data suggest that the H5N1 viruses tested, especially the virus with the high-pathogenicity phenotype, possess greater capability to attenuate the type I IFN response than the human H3N2 virus. The attenuation of this critical host innate immune defense may contribute to the virulence of H5N1 viruses observed in humans.
Figures








References
-
- Chan, M. C., C. Y. Cheung, W. H. Chui, S. W. Tsao, J. M. Nicholls, Y. O. Chan, R. W. Chan, H. T. Long, L. L. Poon, Y. Guan, and J. S. Peiris. 2005. Proinflammatory cytokine responses induced by influenza A (H5N1) viruses in primary human alveolar and bronchial epithelial cells. Respir. Res. 6:135. - PMC - PubMed
-
- Chotpitayasunondh, T., K. Ungchusak, W. Hanshaoworakul, S. Chunsuthiwat, P. Sawanpanyalert, R. Kijphati, S. Lochindarat, P. Srisan, P. Suwan, Y. Osotthanakorn, T. Anantasetagoon, S. Kanjanawasri, S. Tanupattarachai, J. Weerakul, R. Chaiwirattana, M. Maneerattanaporn, R. Poolsavathitikool, K. Chokephaibulkit, A. Apisarnthanarak, and S. F. Dowell. 2005. Human disease from influenza A (H5N1), Thailand, 2004. Emerg. Infect. Dis. 11:201-209. - PMC - PubMed
-
- Coro, E. S., W. L. Chang, and N. Baumgarth. 2006. Type I IFN receptor signals directly stimulate local B cells early following influenza virus infection. J. Immunol. 176:4343-4351. - PubMed
-
- Davies, J. R., S. Kirkham, N. Svitacheva, D. J. Thornton, and I. Carlstedt. 2007. MUC16 is produced in tracheal surface epithelium and submucosal glands and is present in secretions from normal human airway and cultured bronchial epithelial cells. Int. J. Biochem. Cell. Biol. 39:1943-1954. - PubMed
-
- de Jong, M. D., V. C. Bach, T. Q. Phan, M. H. Vo, T. T. Tran, B. H. Nguyen, M. Beld, T. P. Le, H. K. Truong, V. V. Nguyen, T. H. Tran, Q. H. Do, and J. Farrar. 2005. Fatal avian influenza A (H5N1) in a child presenting with diarrhea followed by coma. N. Engl. J. Med. 352:686-691. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
