

Motor force field learning influences visual processing of target motion
- PMID: 17855611
- PMCID: PMC6672632
- DOI: 10.1523/JNEUROSCI.1245-07.2007
Motor force field learning influences visual processing of target motion
Abstract
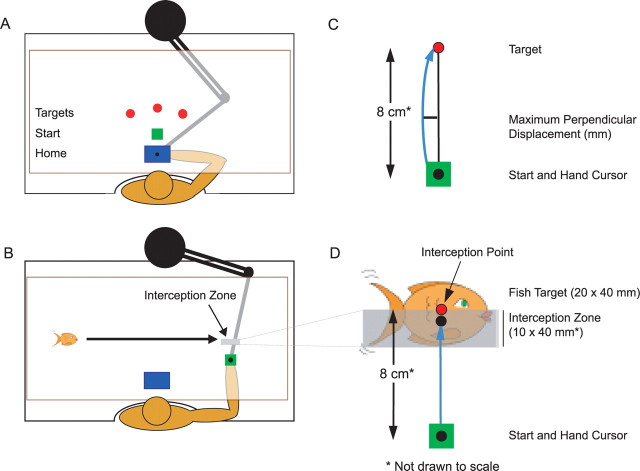
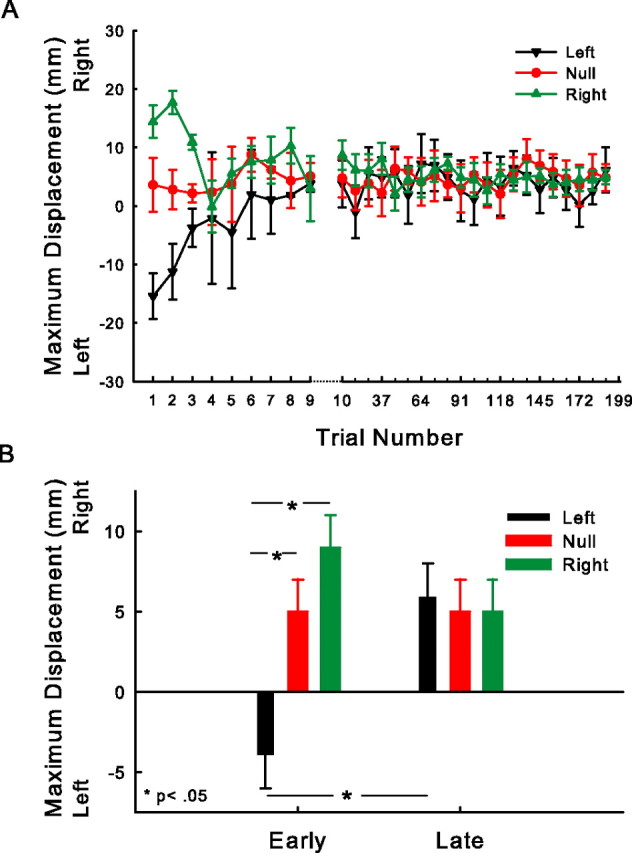
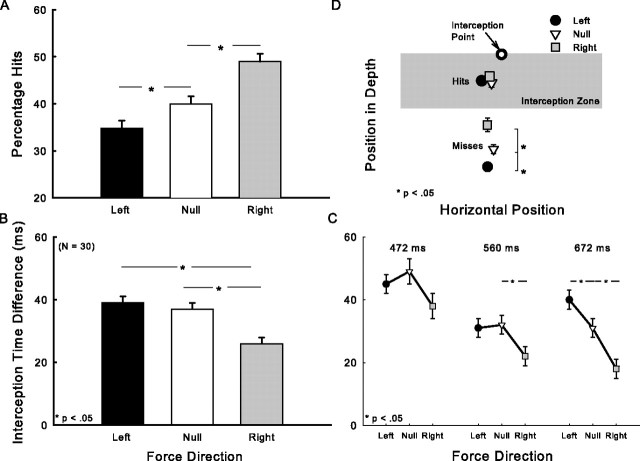
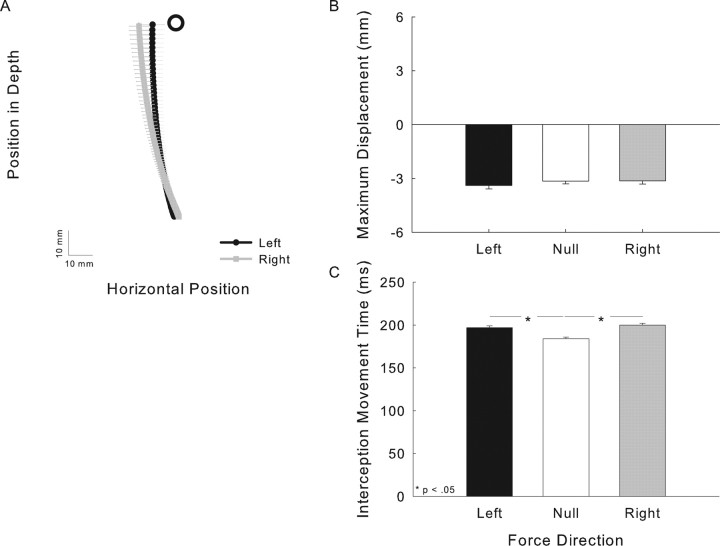
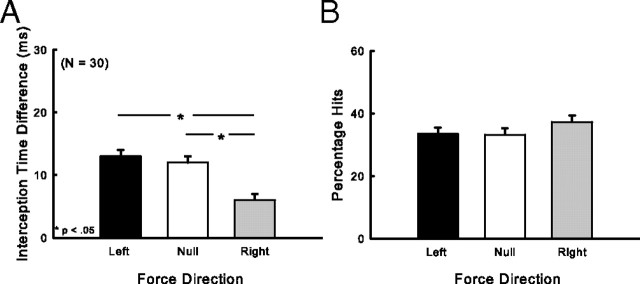
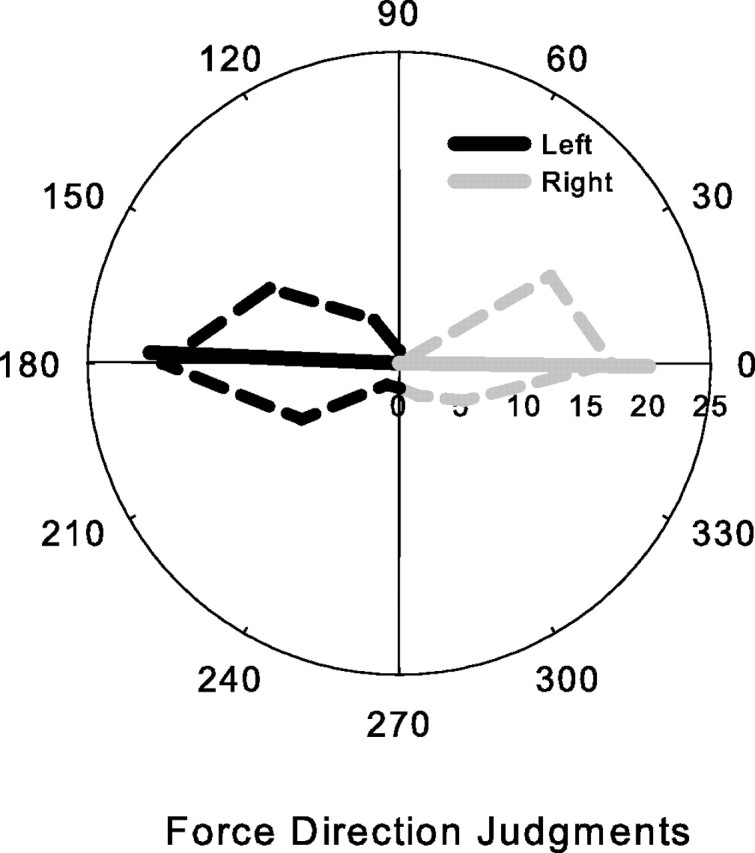
There are reciprocal connections between visual and motor areas of the cerebral cortex. Although recent studies have provided intriguing new insights, in comparison with volume of research on the visual control of movement, relatively little is known about how movement influences vision. The motor system is perfectly suited to learn about environmental forces. Does environmental force information, learned by the motor system, influence visual processing? Here, we show that learning to compensate for a force applied to the hand influenced how participants predicted target motion for interception. Ss trained in one of three constant force fields by making reaching movements while holding a robotic manipulandum. The robot applied forces in a null [null force field (NFF)], leftward [leftward force field (LFF)], or [rightward force field (RFF)] direction. Training was followed immediately with an interception task. The target accelerated from left to right and Ss's task was to stab it. When viewing time was optimal for prediction, the RFF group initiated their responses earlier and hit more targets, and the LFF group initiated their responses later and hit fewer targets, than the NFF group. In follow-up experiments, we show that motor learning is necessary, and we rule out the possibility that explicit force direction information drives how Ss altered their predictions of visual motion. Environmental force information, acquired by motor learning, influenced how the motion of nearby visual targets was predicted.
Figures
Comment in
-
Swimming with and against the stream: does motor adaptation to lateral forces influence visual motion perception?J Neurosci. 2007 Dec 5;27(49):13367-8. doi: 10.1523/JNEUROSCI.4545-07.2007. J Neurosci. 2007. PMID: 18057192 Free PMC article. Review. No abstract available.
Similar articles
-
Controlling reaching movements with predictable and unpredictable target motion in 10-year-old children and adults.Exp Brain Res. 2007 Mar;177(4):483-92. doi: 10.1007/s00221-006-0704-3. Epub 2006 Sep 28. Exp Brain Res. 2007. PMID: 17006685
-
Prior Movement of One Arm Facilitates Motor Adaptation in the Other.J Neurosci. 2023 Jun 7;43(23):4341-4351. doi: 10.1523/JNEUROSCI.2166-22.2023. Epub 2023 May 9. J Neurosci. 2023. PMID: 37160362 Free PMC article.
-
Large-field visual motion directly induces an involuntary rapid manual following response.J Neurosci. 2005 May 18;25(20):4941-51. doi: 10.1523/JNEUROSCI.4143-04.2005. J Neurosci. 2005. PMID: 15901775 Free PMC article.
-
Neurophysiology of perceptual and motor aspects of interception.J Neurophysiol. 2006 Jan;95(1):1-13. doi: 10.1152/jn.00422.2005. J Neurophysiol. 2006. PMID: 16339504 Review.
-
Visual factors in hitting and catching.J Sports Sci. 1997 Dec;15(6):533-58. doi: 10.1080/026404197366985. J Sports Sci. 1997. PMID: 9486432 Review.
Cited by
-
Sensory Plasticity in Human Motor Learning.Trends Neurosci. 2016 Feb;39(2):114-123. doi: 10.1016/j.tins.2015.12.006. Epub 2016 Jan 13. Trends Neurosci. 2016. PMID: 26774345 Free PMC article. Review.
-
Seeing motion of controlled object improves grip timing in adults with autism spectrum condition: evidence for use of inverse dynamics in motor control.Exp Brain Res. 2021 Apr;239(4):1047-1059. doi: 10.1007/s00221-021-06046-3. Epub 2021 Feb 2. Exp Brain Res. 2021. PMID: 33528597
-
Extrapolation of vertical target motion through a brief visual occlusion.Exp Brain Res. 2010 Mar;201(3):365-84. doi: 10.1007/s00221-009-2041-9. Epub 2009 Oct 31. Exp Brain Res. 2010. PMID: 19882150
-
Visuo-motor coordination and internal models for object interception.Exp Brain Res. 2009 Feb;192(4):571-604. doi: 10.1007/s00221-008-1691-3. Epub 2009 Jan 13. Exp Brain Res. 2009. PMID: 19139857 Review.
-
Motor learning and its sensory effects: time course of perceptual change and its presence with gradual introduction of load.J Neurophysiol. 2013 Feb;109(3):782-91. doi: 10.1152/jn.00734.2011. Epub 2012 Nov 7. J Neurophysiol. 2013. PMID: 23136347 Free PMC article.
References
-
- Blake R, Sobel KV, James TW. Neural synergy between kinetic vision and touch. Psychol Sci. 2004;15:397–402. - PubMed
-
- Brashers-Krug T, Shadmehr R, Bizzi E. Consolidation in human motor memory. Nature. 1996;382:252–255. - PubMed
-
- Brouwer AM, Brenner E, Smeets JB. Perception of acceleration with short presentation times: can acceleration be used in interception? Percept Psychophys. 2002;64:1160–1168. - PubMed
-
- Calvo-Merino B, Glaser DE, Grezes J, Passingham RE, Haggard P. Action observation and acquired motor skills: an FMRI study with expert dancers. Cereb Cortex. 2005;15:1243–1249. - PubMed
-
- Casile A, Giese MA. Nonvisual motor training influences biological motion perception. Curr Biol. 2006;16:69–74. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources