

Actomyosin contractility and microtubules drive apical constriction in Xenopus bottle cells
- PMID: 17868669
- PMCID: PMC2744900
- DOI: 10.1016/j.ydbio.2007.08.010
Actomyosin contractility and microtubules drive apical constriction in Xenopus bottle cells
Abstract
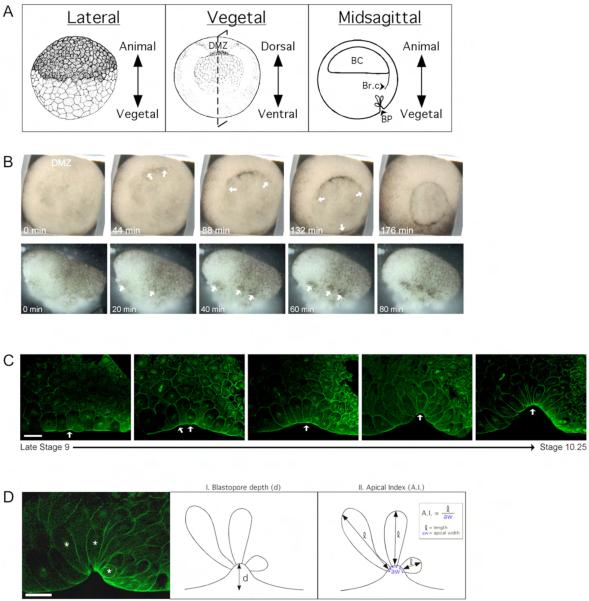
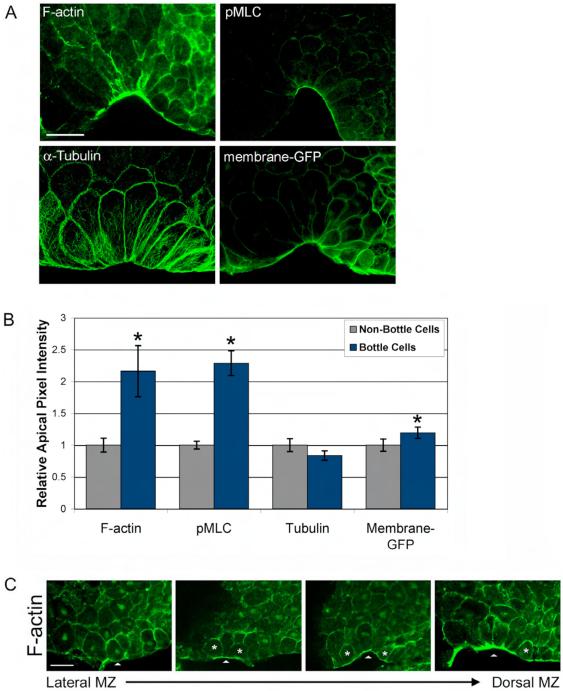
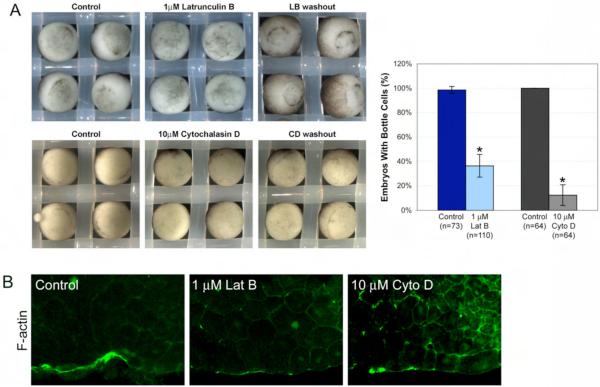
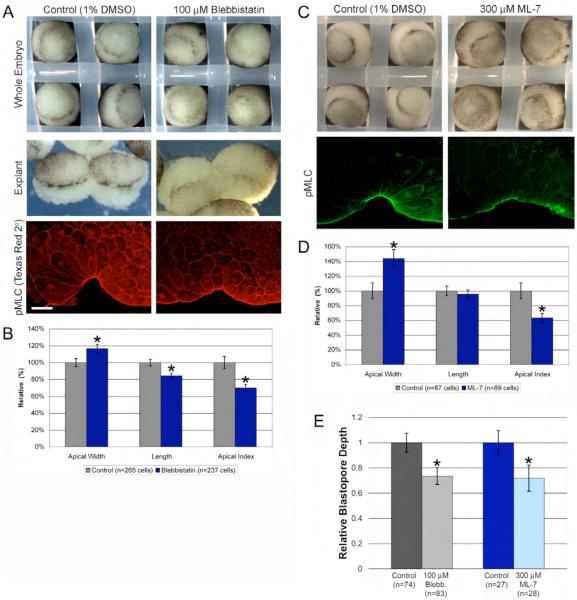
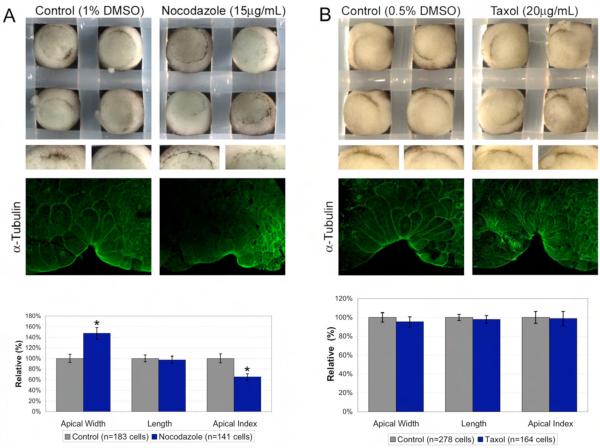
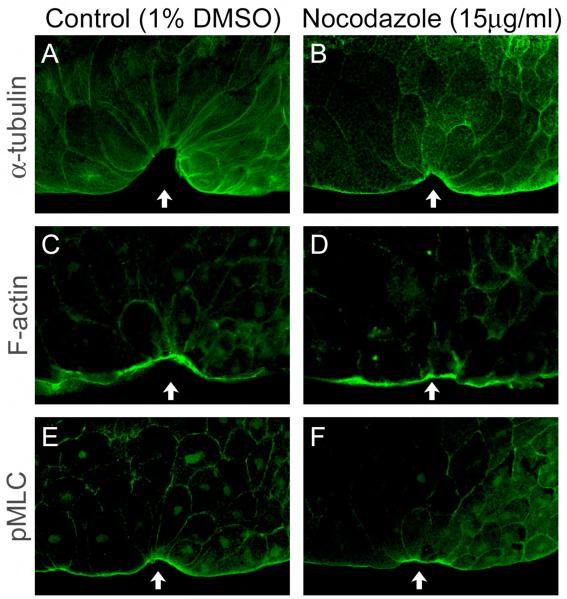
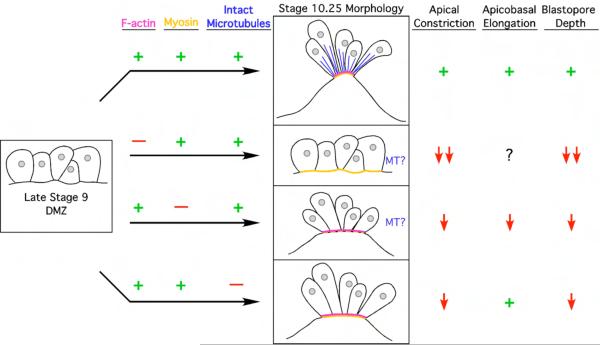
Cell shape changes are critical for morphogenetic events such as gastrulation, neurulation, and organogenesis. However, the cell biology driving cell shape changes is poorly understood, especially in vertebrates. The beginning of Xenopus laevis gastrulation is marked by the apical constriction of bottle cells in the dorsal marginal zone, which bends the tissue and creates a crevice at the blastopore lip. We found that bottle cells contribute significantly to gastrulation, as their shape change can generate the force required for initial blastopore formation. As actin and myosin are often implicated in contraction, we examined their localization and function in bottle cells. F-actin and activated myosin accumulate apically in bottle cells, and actin and myosin inhibitors either prevent or severely perturb bottle cell formation, showing that actomyosin contractility is required for apical constriction. Microtubules were localized in apicobasally directed arrays in bottle cells, emanating from the apical surface. Surprisingly, apical constriction was inhibited in the presence of nocodazole but not taxol, suggesting that intact, but not dynamic, microtubules are required for apical constriction. Our results indicate that actomyosin contractility is required for bottle cell morphogenesis and further suggest a novel and unpredicted role for microtubules during apical constriction.
Figures
Similar articles
-
Apical constriction: themes and variations on a cellular mechanism driving morphogenesis.Development. 2014 May;141(10):1987-98. doi: 10.1242/dev.102228. Development. 2014. PMID: 24803648 Free PMC article. Review.
-
The RhoGEF protein Plekhg5 regulates apical constriction of bottle cells during gastrulation.Development. 2018 Dec 12;145(24):dev168922. doi: 10.1242/dev.168922. Development. 2018. PMID: 30446627 Free PMC article.
-
Uncorking gastrulation: the morphogenetic movement of bottle cells.Wiley Interdiscip Rev Dev Biol. 2012 Mar-Apr;1(2):286-93. doi: 10.1002/wdev.19. Epub 2011 Dec 12. Wiley Interdiscip Rev Dev Biol. 2012. PMID: 23801442 Free PMC article. Review.
-
The RhoGEF protein Plekhg5 regulates medioapical and junctional actomyosin dynamics of apical constriction during Xenopus gastrulation.Mol Biol Cell. 2023 Jun 1;34(7):ar64. doi: 10.1091/mbc.E22-09-0411. Epub 2023 Apr 12. Mol Biol Cell. 2023. PMID: 37043306 Free PMC article.
-
Endocytosis is required for efficient apical constriction during Xenopus gastrulation.Curr Biol. 2010 Feb 9;20(3):253-8. doi: 10.1016/j.cub.2009.12.021. Epub 2010 Jan 21. Curr Biol. 2010. PMID: 20096583 Free PMC article.
Cited by
-
A model of epithelial invagination driven by collective mechanics of identical cells.Biophys J. 2012 Sep 5;103(5):1069-77. doi: 10.1016/j.bpj.2012.07.018. Biophys J. 2012. PMID: 23009857 Free PMC article.
-
Mechanical impact of epithelial-mesenchymal transition on epithelial morphogenesis in Drosophila.Nat Commun. 2019 Jul 4;10(1):2951. doi: 10.1038/s41467-019-10720-0. Nat Commun. 2019. PMID: 31273212 Free PMC article.
-
GEF-H1 functions in apical constriction and cell intercalations and is essential for vertebrate neural tube closure.J Cell Sci. 2014 Jun 1;127(Pt 11):2542-53. doi: 10.1242/jcs.146811. Epub 2014 Mar 28. J Cell Sci. 2014. PMID: 24681784 Free PMC article.
-
Apical constriction: themes and variations on a cellular mechanism driving morphogenesis.Development. 2014 May;141(10):1987-98. doi: 10.1242/dev.102228. Development. 2014. PMID: 24803648 Free PMC article. Review.
-
Stable Force Balance between Epithelial Cells Arises from F-Actin Turnover.Dev Cell. 2015 Dec 21;35(6):685-97. doi: 10.1016/j.devcel.2015.11.018. Epub 2015 Dec 10. Dev Cell. 2015. PMID: 26688336 Free PMC article.
References
-
- Barrett K, Leptin M, Settleman J. The Rho GTPase and a putative RhoGEF mediate a signaling pathway for the cell shape changes in Drosophila gastrulation. Cell. 1997;91:905–15. - PubMed
-
- Bresnick AR. Molecular mechanisms of nonmuscle myosin-II regulation. Curr Opin Cell Biol. 1999;11:26–33. - PubMed
-
- Burnside B. Microtubules and microfilaments in newt neuralation. Dev Biol. 1971;26:416–41. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
