

RNA-specific ribonucleotidyl transferases
- PMID: 17872511
- PMCID: PMC2040100
- DOI: 10.1261/rna.652807
RNA-specific ribonucleotidyl transferases
Abstract
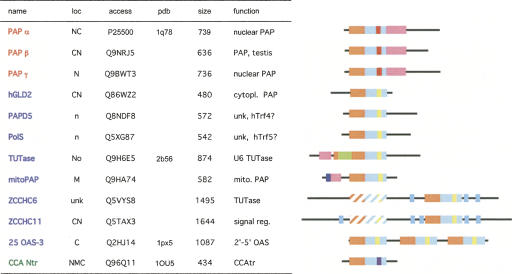
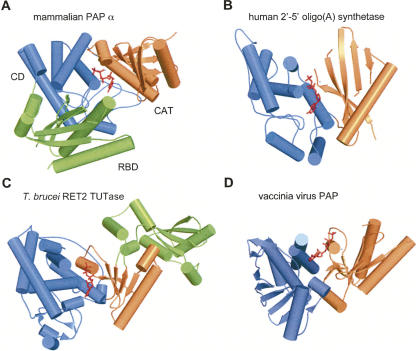
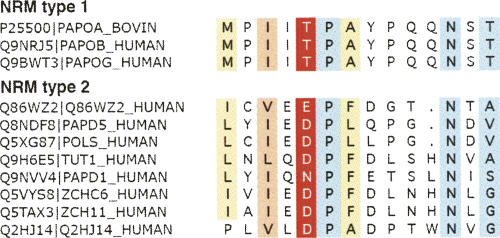
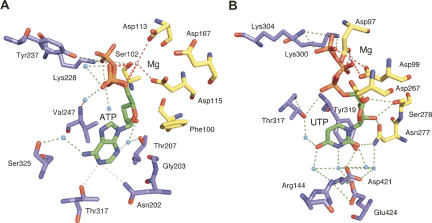
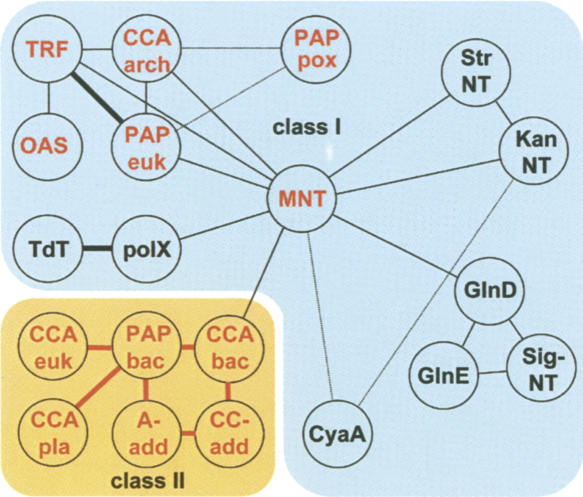
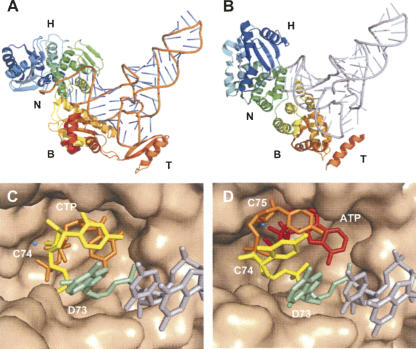
RNA-specific nucleotidyl transferases (rNTrs) are a diverse family of template-independent polymerases that add ribonucleotides to the 3'-ends of RNA molecules. All rNTrs share a related active-site architecture first described for DNA polymerase beta and a catalytic mechanism conserved among DNA and RNA polymerases. The best known examples are the nuclear poly(A) polymerases involved in the 3'-end processing of eukaryotic messenger RNA precursors and the ubiquitous CCA-adding enzymes that complete the 3'-ends of tRNA molecules. In recent years, a growing number of new enzymes have been added to the list that now includes the "noncanonical" poly(A) polymerases involved in RNA quality control or in the readenylation of dormant messenger RNAs in the cytoplasm. Other members of the group are terminal uridylyl transferases adding single or multiple UMP residues in RNA-editing reactions or upon the maturation of small RNAs and poly(U) polymerases, the substrates of which are still not known. 2'-5'Oligo(A) synthetases differ from the other rNTrs by synthesizing oligonucleotides with 2'-5'-phosphodiester bonds de novo.
Figures
Similar articles
-
A family of poly(U) polymerases.RNA. 2007 Jun;13(6):860-7. doi: 10.1261/rna.514007. Epub 2007 Apr 20. RNA. 2007. PMID: 17449726 Free PMC article.
-
Mutational analysis of mammalian poly(A) polymerase identifies a region for primer binding and catalytic domain, homologous to the family X polymerases, and to other nucleotidyltransferases.EMBO J. 1996 May 15;15(10):2593-603. EMBO J. 1996. PMID: 8665867 Free PMC article.
-
Requirement of fission yeast Cid14 in polyadenylation of rRNAs.Mol Cell Biol. 2006 Mar;26(5):1710-21. doi: 10.1128/MCB.26.5.1710-1721.2006. Mol Cell Biol. 2006. PMID: 16478992 Free PMC article.
-
Determinants of substrate specificity in RNA-dependent nucleotidyl transferases.Biochim Biophys Acta. 2008 Apr;1779(4):206-16. doi: 10.1016/j.bbagrm.2007.12.003. Epub 2007 Dec 14. Biochim Biophys Acta. 2008. PMID: 18177750 Free PMC article. Review.
-
Polyadenylation and beyond: emerging roles for noncanonical poly(A) polymerases.Wiley Interdiscip Rev RNA. 2010 Jul-Aug;1(1):142-51. doi: 10.1002/wrna.16. Epub 2010 May 6. Wiley Interdiscip Rev RNA. 2010. PMID: 21956911 Review.
Cited by
-
Inositol 1,4,5-triphosphate receptor-binding protein released with inositol 1,4,5-triphosphate (IRBIT) associates with components of the mRNA 3' processing machinery in a phosphorylation-dependent manner and inhibits polyadenylation.J Biol Chem. 2009 Apr 17;284(16):10694-705. doi: 10.1074/jbc.M807136200. Epub 2009 Feb 18. J Biol Chem. 2009. PMID: 19224921 Free PMC article.
-
The 'how' and 'where' of plant microRNAs.New Phytol. 2017 Dec;216(4):1002-1017. doi: 10.1111/nph.14834. Epub 2017 Oct 19. New Phytol. 2017. PMID: 29048752 Free PMC article. Review.
-
Role of RNA modifications in cancer.Nat Rev Cancer. 2020 Jun;20(6):303-322. doi: 10.1038/s41568-020-0253-2. Epub 2020 Apr 16. Nat Rev Cancer. 2020. PMID: 32300195 Review.
-
Nontemplate-driven polymers: clues to a minimal form of organization closure at the early stages of living systems.Theory Biosci. 2015 Jun;134(1-2):47-64. doi: 10.1007/s12064-015-0209-3. Epub 2015 Apr 28. Theory Biosci. 2015. PMID: 25916275
-
Widespread RNA 3'-end oligouridylation in mammals.RNA. 2012 Mar;18(3):394-401. doi: 10.1261/rna.029306.111. Epub 2012 Jan 30. RNA. 2012. PMID: 22291204 Free PMC article.
References
-
- Aphasizhev, R., Aphasizheva, I., Simpson, L. Multiple terminal uridylyltransferases of trypanosomes. FEBS Lett. 2004;572:15–18. - PubMed
-
- August, J.T., Ortiz, P.J., Hurwitz, J. Ribonucleic acid-dependent ribonucleotide incorporation. I. Purification and properties of the enzyme. J. Biol. Chem. 1962;237:3786–3793. - PubMed
-
- Augustin, M.A., Reichert, A.S., Betat, H., Huber, R., Mörl, M., Steegborn, C. Crystal structure of the human CCA-adding enzyme: Insights into template-independent polymerization. J. Mol. Biol. 2003;328:985–994. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases