

Region-specific network plasticity in simulated and living cortical networks: comparison of the center of activity trajectory (CAT) with other statistics
- PMID: 17873432
- PMCID: PMC2577565
- DOI: 10.1088/1741-2560/4/3/015
Region-specific network plasticity in simulated and living cortical networks: comparison of the center of activity trajectory (CAT) with other statistics
Abstract
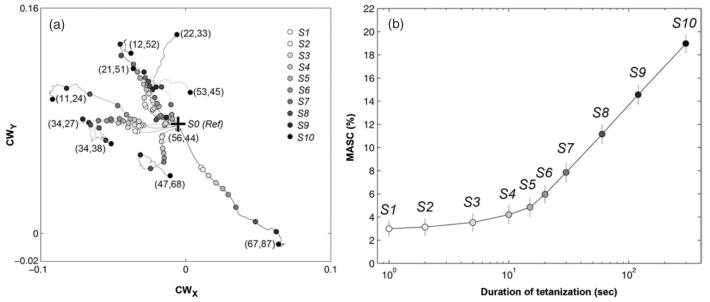
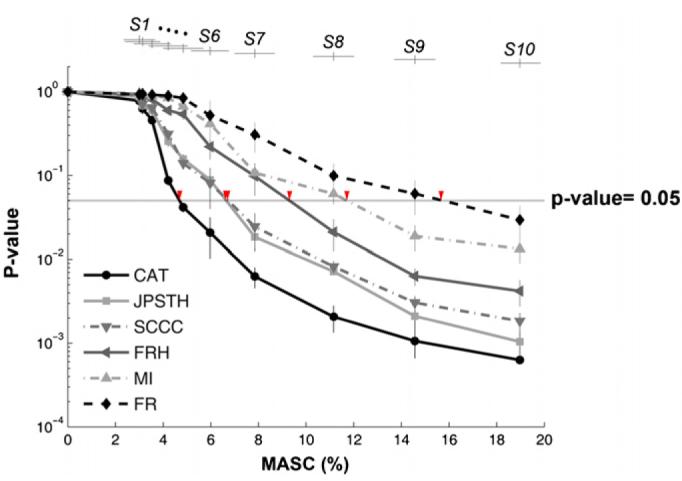
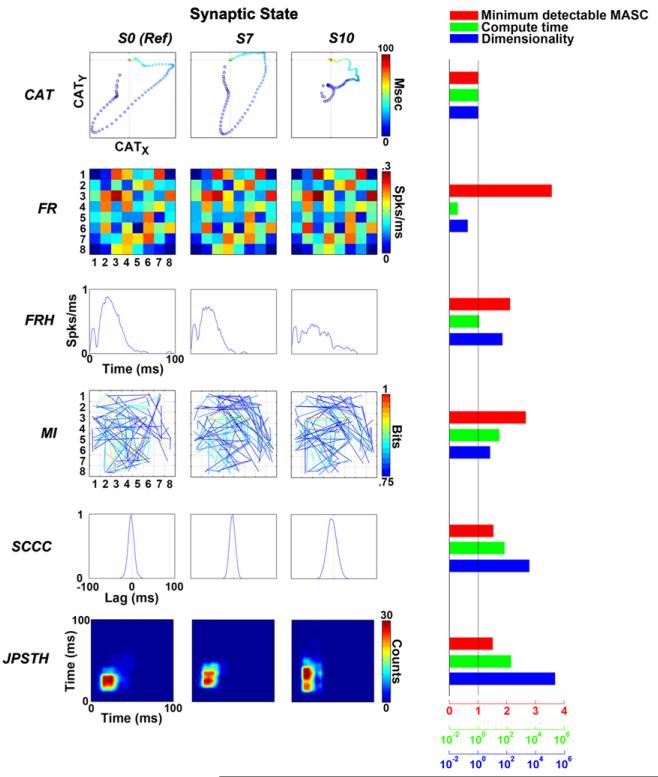
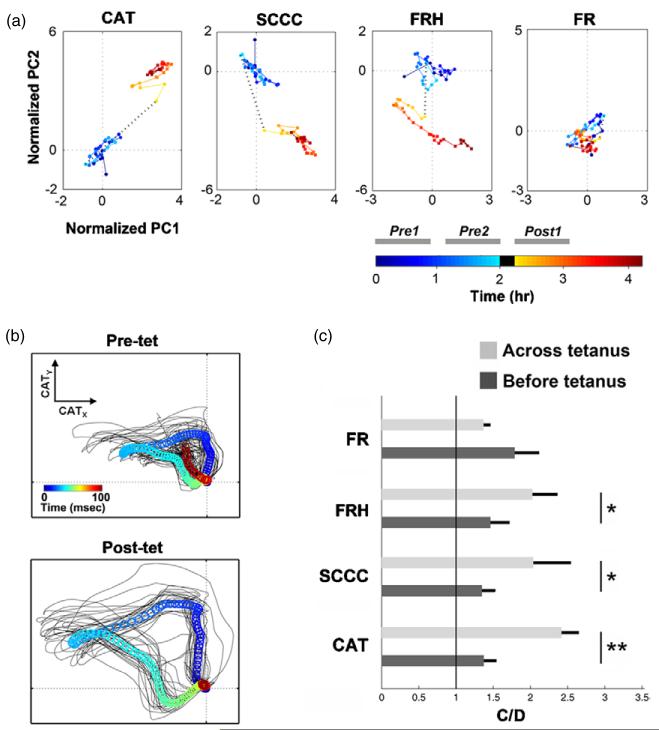
Electrically interfaced cortical networks cultured in vitro can be used as a model for studying the network mechanisms of learning and memory. Lasting changes in functional connectivity have been difficult to detect with extracellular multi-electrode arrays using standard firing rate statistics. We used both simulated and living networks to compare the ability of various statistics to quantify functional plasticity at the network level. Using a simulated integrate-and-fire neural network, we compared five established statistical methods to one of our own design, called center of activity trajectory (CAT). CAT, which depicts dynamics of the location-weighted average of spatiotemporal patterns of action potentials across the physical space of the neuronal circuitry, was the most sensitive statistic for detecting tetanus-induced plasticity in both simulated and living networks. By reducing the dimensionality of multi-unit data while still including spatial information, CAT allows efficient real-time computation of spatiotemporal activity patterns. Thus, CAT will be useful for studies in vivo or in vitro in which the locations of recording sites on multi-electrode probes are important.
Figures
Similar articles
-
Understanding the temporal evolution of neuronal connectivity in cultured networks using statistical analysis.BMC Neurosci. 2014 Jan 21;15:17. doi: 10.1186/1471-2202-15-17. BMC Neurosci. 2014. PMID: 24443925 Free PMC article.
-
Effects of random external background stimulation on network synaptic stability after tetanization: a modeling study.Neuroinformatics. 2005;3(3):263-80. doi: 10.1385/NI:3:3:263. Neuroinformatics. 2005. PMID: 16077162 Free PMC article.
-
Do external stimuli, applied to train cultured cortical networks, disturb the balance between activity and connectivity?Annu Int Conf IEEE Eng Med Biol Soc. 2008;2008:5081-4. doi: 10.1109/IEMBS.2008.4650356. Annu Int Conf IEEE Eng Med Biol Soc. 2008. PMID: 19163859
-
Plasticity and specificity of cortical processing networks.Trends Neurosci. 2006 Jun;29(6):323-9. doi: 10.1016/j.tins.2006.04.002. Epub 2006 May 11. Trends Neurosci. 2006. PMID: 16697057 Review.
-
Spontaneous and driven cortical activity: implications for computation.Curr Opin Neurobiol. 2009 Aug;19(4):439-44. doi: 10.1016/j.conb.2009.07.005. Epub 2009 Aug 3. Curr Opin Neurobiol. 2009. PMID: 19647992 Free PMC article. Review.
Cited by
-
Understanding the temporal evolution of neuronal connectivity in cultured networks using statistical analysis.BMC Neurosci. 2014 Jan 21;15:17. doi: 10.1186/1471-2202-15-17. BMC Neurosci. 2014. PMID: 24443925 Free PMC article.
-
Plasticity of recurring spatiotemporal activity patterns in cortical networks.Phys Biol. 2007 Oct 9;4(3):181-93. doi: 10.1088/1478-3975/4/3/005. Phys Biol. 2007. PMID: 17928657 Free PMC article.
-
Spatial Memory in a Spiking Neural Network with Robot Embodiment.Sensors (Basel). 2021 Apr 10;21(8):2678. doi: 10.3390/s21082678. Sensors (Basel). 2021. PMID: 33920246 Free PMC article.
-
MEART: The Semi-Living Artist.Front Neurorobot. 2007 Nov 2;1:5. doi: 10.3389/neuro.12.005.2007. eCollection 2007. Front Neurorobot. 2007. PMID: 18958276 Free PMC article.
-
Controlling neural network responsiveness: tradeoffs and constraints.Front Neuroeng. 2014 Apr 29;7:11. doi: 10.3389/fneng.2014.00011. eCollection 2014. Front Neuroeng. 2014. PMID: 24808860 Free PMC article.
References
-
- Ahissar E, Abeles M, Ahissar M, Haidarliu S, Vaadia E. Hebbian-like functional plasticity in the auditory cortex of the behaving monkey. Neuropharmacology. 1998;37:633–55. - PubMed
-
- Ahissar E, Vaadia E, Ahissar M, Bergman H, Arieli A, Abeles M. Dependence of cortical plasticity on correlated activity of single neurons and on behavioral context. Science. 1992;257:1412–5. - PubMed
-
- Baruchi I, Ben-Jacob E. Functional holography of recorded neuronal networks activity. J. Neuroinform. 2004;2:333–52. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous