

Melanopsin-dependent nonvisual responses: evidence for photopigment bistability in vivo
- PMID: 17876062
- PMCID: PMC2789279
- DOI: 10.1177/0748730407306043
Melanopsin-dependent nonvisual responses: evidence for photopigment bistability in vivo
Abstract
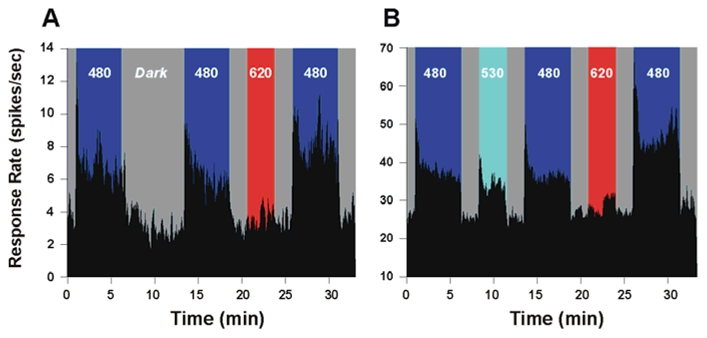
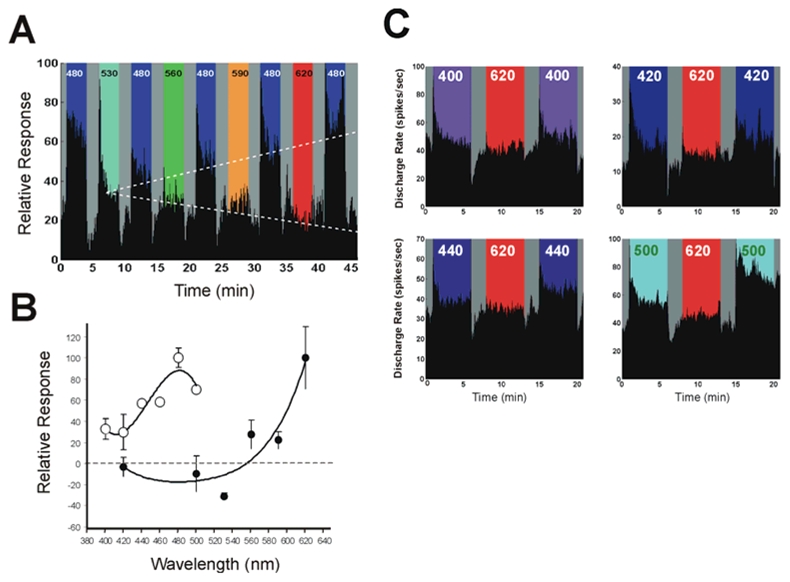
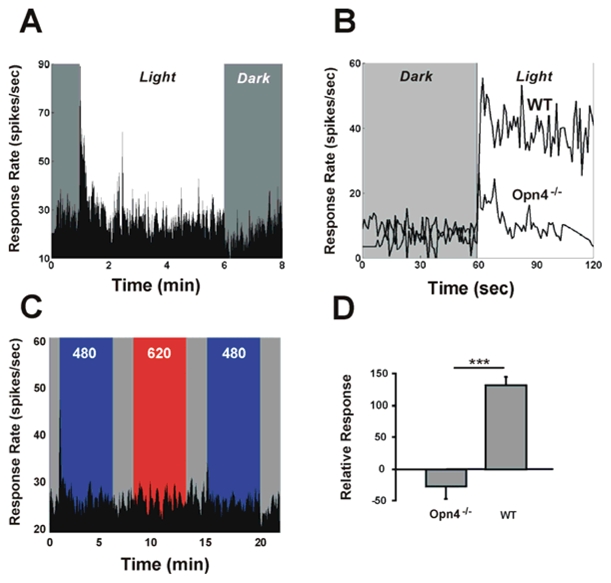
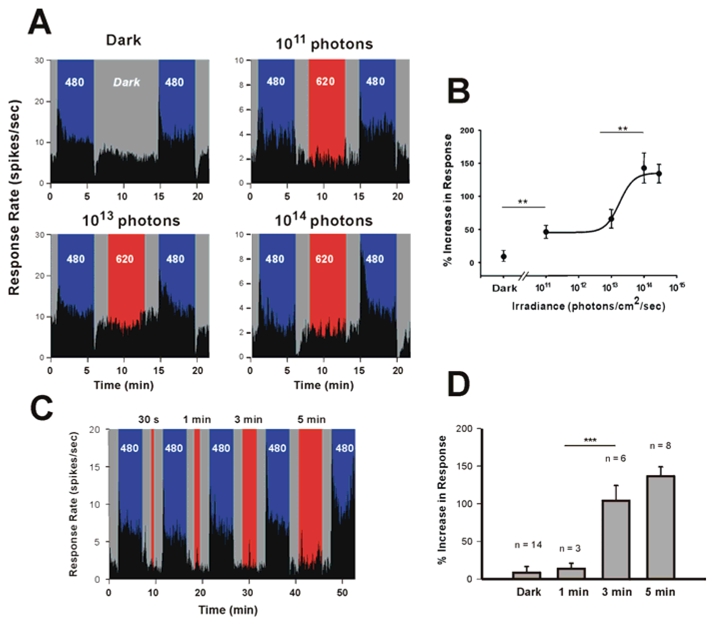
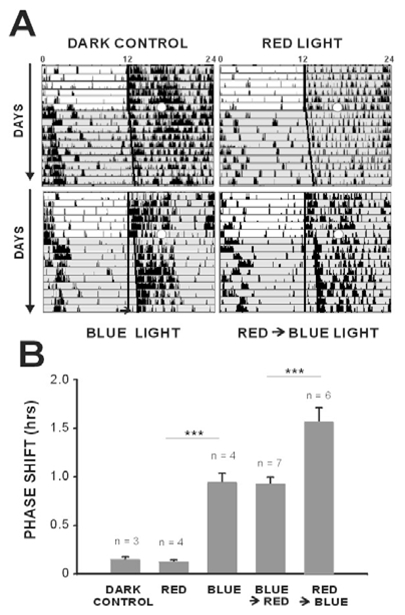
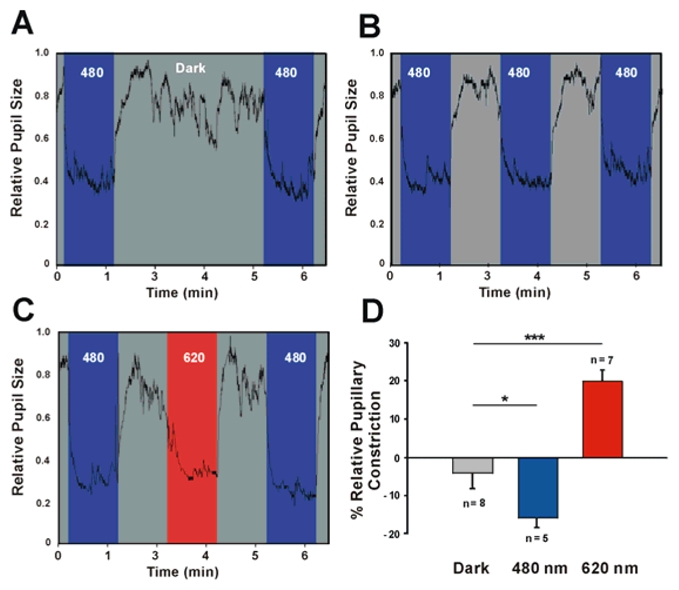
In mammals, nonvisual responses to light have been shown to involve intrinsically photosensitive retinal ganglion cells (ipRGC) that express melanopsin and that are modulated by input from both rods and cones. Recent in vitro evidence suggests that melanopsin possesses dual photosensory and photoisomerase functions, previously thought to be a unique feature of invertebrate rhabdomeric photopigments. In cultured cells that normally do not respond to light, heterologous expression of mammalian melanopsin confers light sensitivity that can be restored by prior stimulation with appropriate wavelengths. Using three different physiological and behavioral assays, we show that this in vitro property translates to in vivo, melanopsin-dependent nonvisual responses. We find that prestimulation with long-wavelength light not only restores but enhances single-unit responses of SCN neurons to 480-nm light, whereas the long-wavelength stimulus alone fails to elicit any response. Recordings in Opn4-/- mice confirm that melanopsin provides the main photosensory input to the SCN, and furthermore, demonstrate that melanopsin is required for response enhancement, because this capacity is abolished in the knockout mouse. The efficiency of the light-enhancement effect depends on wavelength, irradiance, and duration. Prior long-wavelength light exposure also enhances short-wavelength-induced phase shifts of locomotor activity and pupillary constriction, consistent with the expression of a photoisomerase-like function in nonvisual responses to light.
Figures
Comment in
-
Does melanopsin bistability have physiological consequences?J Biol Rhythms. 2008 Oct;23(5):396-9. doi: 10.1177/0748730408323067. J Biol Rhythms. 2008. PMID: 18838605
References
-
- Arendt D. Evolution of eyes and photoreceptor cell types. Int J Dev Biol. 2003;47:563–571. - PubMed
-
- Arendt D, Tessmar-Raible K, Snyman H, Dorresteijn AW, Wittbrodt J. Ciliary photoreceptors with a vertebrate-type opsin in an invertebrate brain. Science. 2004;306:869–871. - PubMed
-
- Arshavsky V. Like night and day: rods and cones have different pigment regeneration pathways. Neuron. 2002;36:1–3. - PubMed
-
- Belenky MA, Smeraski CA, Provencio I, Sollars PJ, Pickard GE. Melanopsin retinal ganglion cells receive bipolar and amacrine cell synapses. J Comp Neurol. 2003;460:380–393. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
