

Cell cycle-dependent expression of potassium channels and cell proliferation in rat mesenchymal stem cells from bone marrow
- PMID: 17877608
- PMCID: PMC6496559
- DOI: 10.1111/j.1365-2184.2007.00458.x
Cell cycle-dependent expression of potassium channels and cell proliferation in rat mesenchymal stem cells from bone marrow
Abstract
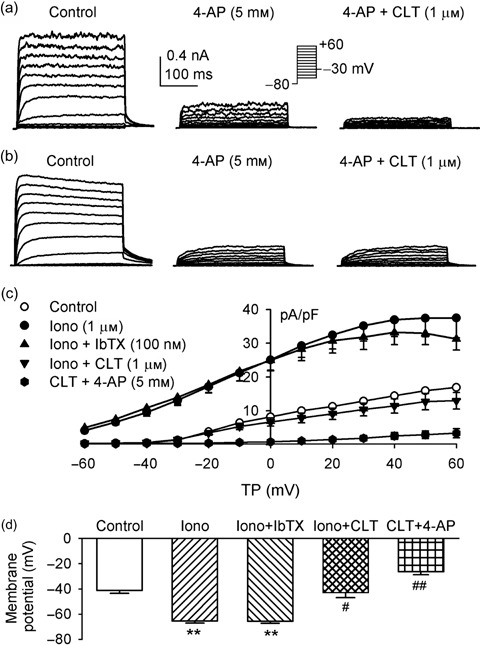
Objective: Recently, our team has demonstrated that voltage-gated delayed rectifier K(+) current (IK(DR)) and Ca(2+)-activated K(+) current (I(KCa)) are present in rat bone marrow-derived mesenchymal stem cells; however, little is known of their physiological roles. The present study was designed to investigate whether functional expression of IK(DR) and I(KCa) would change with cell cycle progression, and whether they could regulate proliferation in undifferentiated rat mesenchymal stem cells (MSCs).
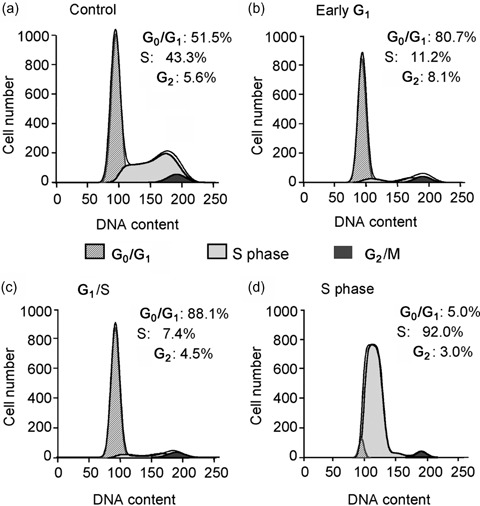
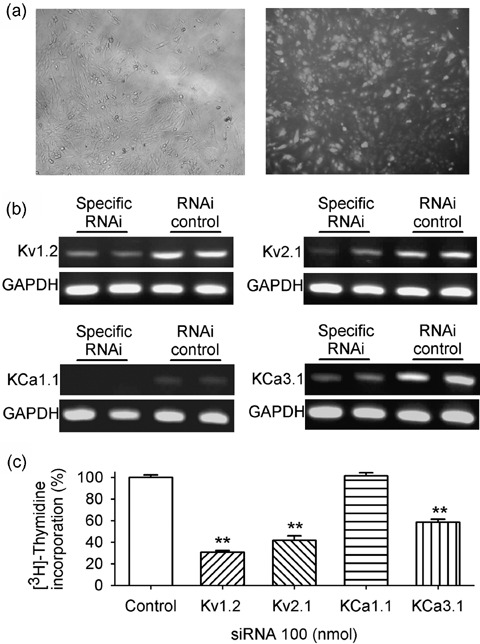
Materials and methods: Membrane potentials and ionic currents were recorded using whole-cell patch clamp technique, cell cycling was analysed by flow cytometry, cell proliferation was assayed with DNA incorporation method and the related genes were down-regulated by RNA interference (RNAi) and examined using RT-PCR.
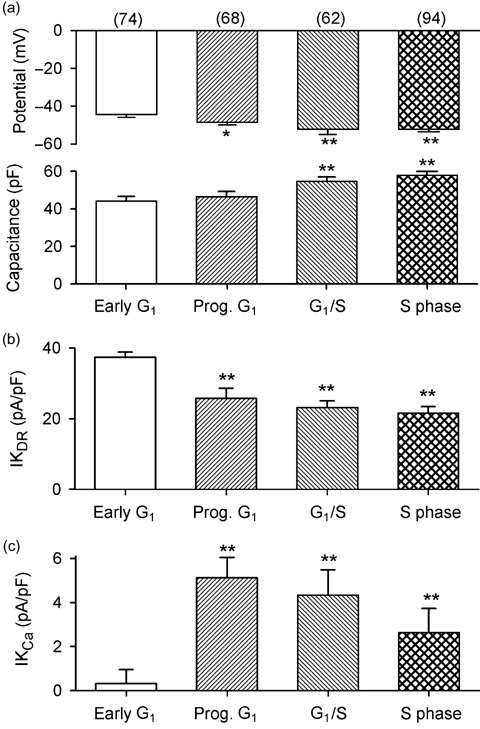
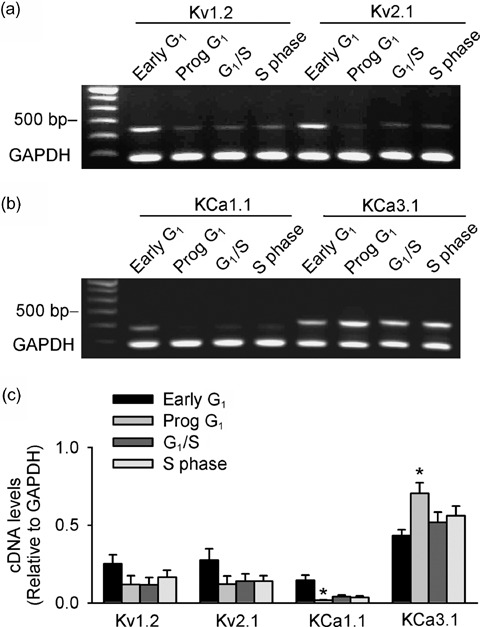
Results: It was found that membrane potential hyperpolarized, and cell size increased during the cell cycle. In addition, IK(DR) decreased, while I(KCa) increased during progress from G(1) to S phase. RT-PCR revealed that the mRNA levels of Kv1.2 and Kv2.1 (likely responsible for IK(DR)) reduced, whereas the mRNA level of KCa3.1 (responsible for intermediate-conductance I(KCa)) increased with the cell cycle progression. Down-regulation of Kv1.2, Kv2.1 or KCa3.1 with the specific RNAi, targeted to corresponding gene inhibited proliferation of rat MSCs.
Conclusion: These results demonstrate that membrane potential, IK(DR) and I(KCa) channels change with cell cycle progression and corresponding alteration of gene expression. IK(DR) and intermediate-conductance I(KCa) play an important role in maintaining membrane potential and they participate in modulation of proliferation in rat MSCs.
Figures
References
-
- Bruder SP, Jaiswal N, Haynesworth SE (1997) Growth kinetics, self‐renewal, and the osteogenic potential of purified human mesenchymal stem cells during extensive subcultivation and following cryopreservation. J. Cell. Biochem. 64, 278–294. - PubMed
-
- Cahalan MD, Wulff H, Chandy KG (2001) Molecular properties and physiological roles of ion channels in the immune system. J. Clin. Immunol. 21, 235–252. - PubMed
-
- Cahill KS, Toma C, Pittenger MF, Kessler PD, Byrne BJ (2003) Cell therapy in the heart: cell production, transplantation, and applications. Methods Mol. Biol. 219, 73–81. - PubMed
-
- Caplan AI, Bruder SP (2001) Mesenchymal stem cells: building blocks for molecular medicine in the 21st century. Trends Mol. Med. 7, 259–264. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
