

Cyclin-dependent kinase inhibitors and basement membrane interact to regulate breast epithelial cell differentiation and acinar morphogenesis
- PMID: 17877612
- PMCID: PMC6496798
- DOI: 10.1111/j.1365-2184.2007.00463.x
Cyclin-dependent kinase inhibitors and basement membrane interact to regulate breast epithelial cell differentiation and acinar morphogenesis
Abstract
Objective: The cyclin-dependent kinase inhibitors (CDKIs), p21(CIP1) and p27(KIP1) regulate growth and differentiation in diverse tissue types. We aimed to determine whether p21(CIP1) or p27(KIP1) could induce a terminally differentiated phenotype in breast cells, and to examine if CDKI expression is regulated by basement membrane interactions.
Materials and methods: Effects of increased CDKI expression on the phenotype of MCF-10A breast epithelial cells were examined by retroviral transduction of p21(CIP1) or p27(KIP1) cDNA.
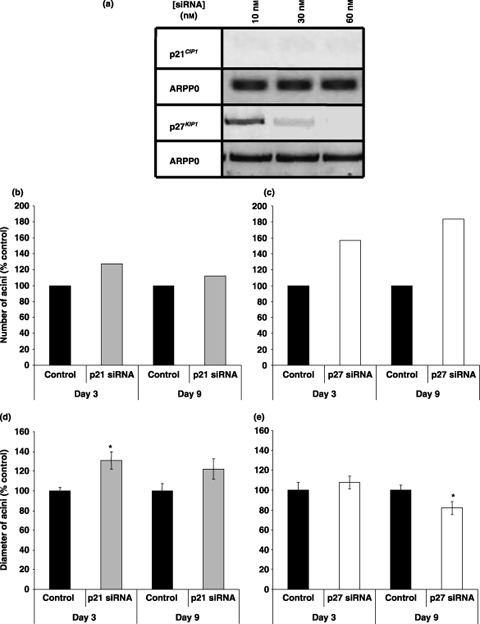
Results: Overexpression of p21(CIP1) or p27(KIP1) reduced MCF-10A growth rates in monolayer cultures, altered cellular morphology and stimulated accumulation of neutral lipid droplets, suggesting partial lactational differentiation. However, markers of luminal differentiation (oestrogen and progesterone receptors, alpha-lactalbumin, beta-casein and adipophilin) were absent when examined by reverse transcriptase-polymerase chain reaction and immunohistochemistry. Cell-basement membrane contacts are known to be essential for full mammary epithelial cell differentiation and therefore parental MCF-10A cells were cultured on a basement membrane preparation (Matrigel) in which they form acini. Immunocytochemistry showed that Ki67, the cell proliferation marker, was initially expressed at high levels and as growth decreased p27(KIP1) expression steadily increased. Surprisingly, p21(CIP1) was highest at the early stages of acinus growth and was detected in proliferating cells, as demonstrated by colocalization in dual Ki67/p21(CIP1) immunofluorescence. Overexpression of p21(CIP1) or p27(KIP1) impaired formation of acini, whereas their knockdown, using siRNA, increased acinus formation.
Conclusion: We conclude that both p21(CIP1) and p27(KIP1) induce partial secretory differentiation of mammary cells in monolayer, but during acinus morphogenesis in 3D culture they have a highly regulated temporal expression pattern.
Figures
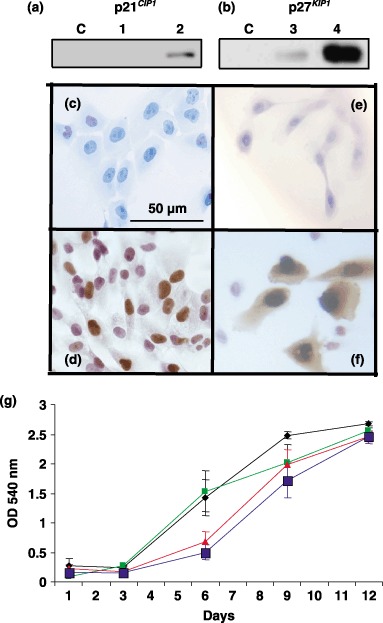
 ) and p27KIP1 (
) and p27KIP1 ( ) transduced cells showed retarded growth compared to control cells (
) transduced cells showed retarded growth compared to control cells ( ) or vector treated cells (). Results are expressed as average ± SEM.
) or vector treated cells (). Results are expressed as average ± SEM.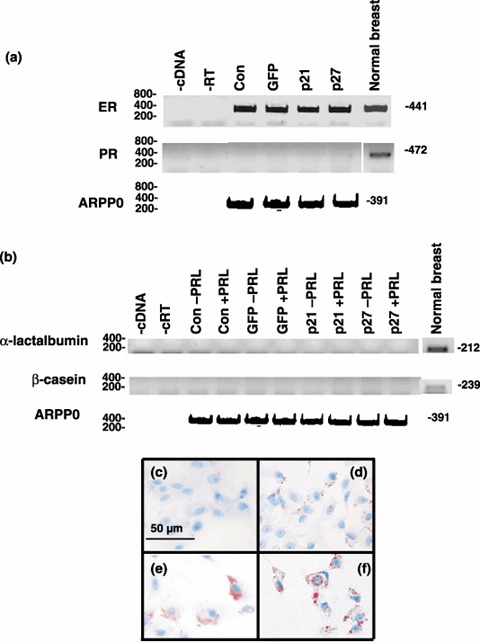
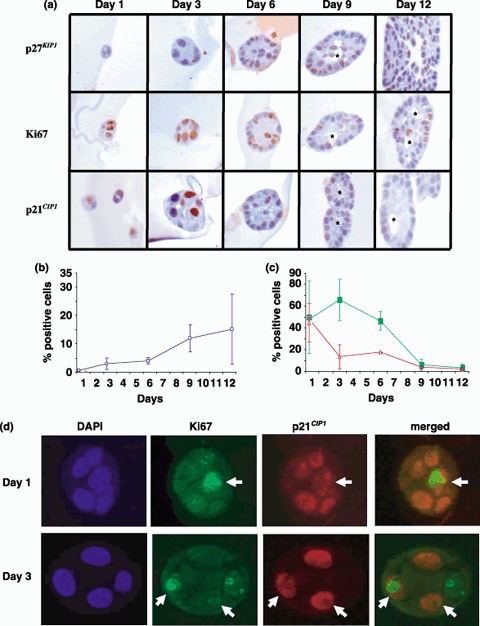 ) (b), Ki67 () and p21CIP1 () (c). (d) Representative photomicrographs showing colocalization of Ki67 and p21CIP1 MCF‐10A cells were cultured on Matrigel, fixed and sectioned as described in Materials and Methods. Sections were subjected to dual immunofluorescence for p21CIP1 (red) and Ki67 (green). Nuclei were counterstained with DAPI (blue). Dual‐labelled cells are indicated by arrows.
) (b), Ki67 () and p21CIP1 () (c). (d) Representative photomicrographs showing colocalization of Ki67 and p21CIP1 MCF‐10A cells were cultured on Matrigel, fixed and sectioned as described in Materials and Methods. Sections were subjected to dual immunofluorescence for p21CIP1 (red) and Ki67 (green). Nuclei were counterstained with DAPI (blue). Dual‐labelled cells are indicated by arrows.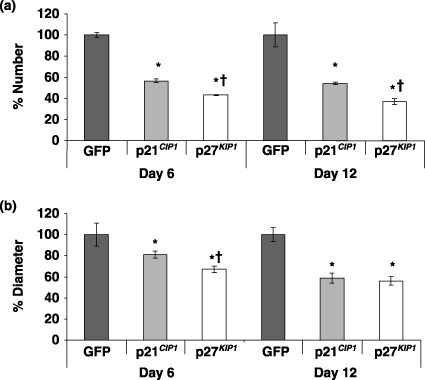
Similar articles
-
Cortactin modulates RhoA activation and expression of Cip/Kip cyclin-dependent kinase inhibitors to promote cell cycle progression in 11q13-amplified head and neck squamous cell carcinoma cells.Mol Cell Biol. 2010 Nov;30(21):5057-70. doi: 10.1128/MCB.00249-10. Epub 2010 Aug 30. Mol Cell Biol. 2010. PMID: 20805359 Free PMC article.
-
p21Cip1 and p27Kip1 induce distinct cell cycle effects and differentiation programs in myeloid leukemia cells.J Biol Chem. 2005 May 6;280(18):18120-9. doi: 10.1074/jbc.M500758200. Epub 2005 Mar 3. J Biol Chem. 2005. PMID: 15746092
-
Expression patterns of cyclins D1, E and cyclin-dependent kinase inhibitors p21(Waf1/Cip1) and p27(Kip1) in urothelial carcinoma: correlation with other cell-cycle-related proteins (Rb, p53, Ki-67 and PCNA) and clinicopathological features.Urol Int. 2004;73(1):65-73. doi: 10.1159/000078807. Urol Int. 2004. PMID: 15263796
-
Ubiquitylation and proteasomal degradation of the p21(Cip1), p27(Kip1) and p57(Kip2) CDK inhibitors.Cell Cycle. 2010 Jun 15;9(12):2342-52. doi: 10.4161/cc.9.12.11988. Epub 2010 Jun 15. Cell Cycle. 2010. PMID: 20519948 Free PMC article. Review.
-
Multiple degradation pathways regulate versatile CIP/KIP CDK inhibitors.Trends Cell Biol. 2012 Jan;22(1):33-41. doi: 10.1016/j.tcb.2011.10.004. Epub 2011 Dec 9. Trends Cell Biol. 2012. PMID: 22154077 Free PMC article. Review.
Cited by
-
Integrin β1 subunit regulates cellular and secreted MUC5AC and MUC5B production in NCI-H292 human lung epithelial cells.Biochem Biophys Rep. 2021 Sep 1;28:101124. doi: 10.1016/j.bbrep.2021.101124. eCollection 2021 Dec. Biochem Biophys Rep. 2021. PMID: 34504957 Free PMC article.
-
Atbf1 regulates pubertal mammary gland development likely by inhibiting the pro-proliferative function of estrogen-ER signaling.PLoS One. 2012;7(12):e51283. doi: 10.1371/journal.pone.0051283. Epub 2012 Dec 12. PLoS One. 2012. PMID: 23251482 Free PMC article.
-
Type IV collagen reduces mucin 5AC secretion in three-dimensional cultured human primary airway epithelial cells.Biochem Biophys Rep. 2019 Nov 8;20:100707. doi: 10.1016/j.bbrep.2019.100707. eCollection 2019 Dec. Biochem Biophys Rep. 2019. PMID: 31737795 Free PMC article.
-
A truncated progesterone receptor (PR-M) localizes to the mitochondrion and controls cellular respiration.Mol Endocrinol. 2013 May;27(5):741-53. doi: 10.1210/me.2012-1292. Epub 2013 Mar 21. Mol Endocrinol. 2013. PMID: 23518922 Free PMC article.
-
Type IV collagen reduces MUC5AC secretion in the lungs of ovalbumin-sensitized mice.Front Pharmacol. 2022 Sep 16;13:851374. doi: 10.3389/fphar.2022.851374. eCollection 2022. Front Pharmacol. 2022. PMID: 36188610 Free PMC article.
References
-
- Aggeler J, Ward J, Blackie LM, Barcellos‐Hoff MH, Streuli CH, Bissell MJ (1991) Cytodifferentiation of mouse mammary epithelial cells cultured on a reconstituted basement membrane reveals striking similarities to development in vivo . J. Cell Sci. 99, 407–417. - PubMed
-
- Anderson E, Clarke RB, Howell A (1998) Estrogen responsiveness and control of normal human breast proliferation. J. Mammary Gland Biol. Neoplasia 3, 23–35. - PubMed
-
- Buckley S, Driscoll B, Anderson KD, Warburton D (1997) Cell cycle in alveolar epithelial type II cells: integration of Matrigel and KGF. Am. J. Physiol. 273, L572–L580. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
