

Activation by insulin and amino acids of signaling components leading to translation initiation in skeletal muscle of neonatal pigs is developmentally regulated
- PMID: 17878222
- PMCID: PMC2714663
- DOI: 10.1152/ajpendo.00307.2007
Activation by insulin and amino acids of signaling components leading to translation initiation in skeletal muscle of neonatal pigs is developmentally regulated
Abstract
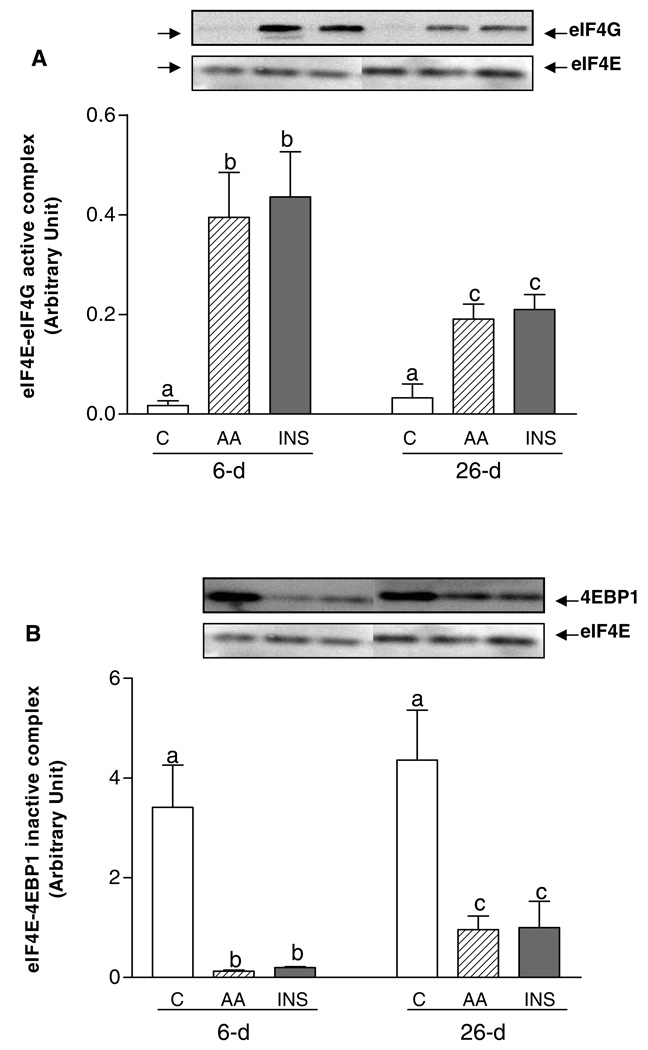
Insulin and amino acids act independently to stimulate protein synthesis in skeletal muscle of neonatal pigs, and the responses decrease with development. The purpose of this study was to compare the separate effects of fed levels of INS and AA on the activation of signaling components leading to translation initiation and how these responses change with development. Overnight-fasted 6- (n = 4/group) and 26-day-old (n = 6/ group) pigs were studied during 1) euinsulinemic-euglycemiceuaminoacidemic conditions (controls), 2) euinsulinemic-euglycemichyperaminoacidemic clamps (AA), and 3) hyperinsulinemic-euglycemic-euaminoacidemic clamps (INS). INS, but not AA, increased the phosphorylation of protein kinase B (PKB) and tuberous sclerosis 2 (TSC2). Both INS and AA increased protein synthesis and the phosphorylation of mammalian target of rapamycin (mTOR), ribosomal protein S6 kinase-1, and eukaryotic initiation factor (eIF)4E-binding protein 1 (4E-BP1), and these responses were higher in 6-day-old compared with 26-day-old pigs. Both INS and AA decreased the binding of 4E-BP1 to eIF4E and increased eIF4E binding to eIF4G; these effects were greater in 6-day-old than in 26-day-old pigs. Neither INS nor AA altered the composition of mTORC1 (raptor, mTOR, and GbetaL) or mTORC2 (rictor, mTOR, and GbetaL) complexes. Furthermore, neither INS, AA, nor age had any effect on the abundance of Rheb and the phosphorylation of AMP-activated protein kinase and eukaryotic elongation factor 2. Our results suggest that the activation by insulin and amino acids of signaling components leading to translation initiation is developmentally regulated and parallels the developmental decline in protein synthesis in skeletal muscle of neonatal pigs.
Figures





References
-
- Asnaghi L, Bruno P, Priulla M, Nicolin A. mTOR: a protein kinase switching between life and death. Pharmacol Res. 2004;50:545–549. - PubMed
-
- Avruch J, Hara K, Lin Y, Liu M, Long X, Ortiz-Vega S, Yonezawa K. Insulin and amino-acid regulation of mTOR signaling and kinase activity through the Rheb GTPase. Oncogene. 2006;25:6361–6372. - PubMed
-
- Avruch J, Lin Y, Long X, Murthy S, Ortiz-Vega S. Recent advances in the regulation of the TOR pathway by insulin and amino acids. Curr Opin Clin Nutr Metab Care. 2005;8:67–72. - PubMed
-
- Bevan P. Insulin signalling. J Cell Sci. 2001;114:1429–1430. - PubMed
-
- Carling D. AMP-activated protein kinase: balancing the scales. Biochimie. 2005;87:87–91. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous