

The IkappaB kinase regulates chromatin structure during reconsolidation of conditioned fear memories
- PMID: 17880897
- PMCID: PMC2587178
- DOI: 10.1016/j.neuron.2007.07.039
The IkappaB kinase regulates chromatin structure during reconsolidation of conditioned fear memories
Abstract
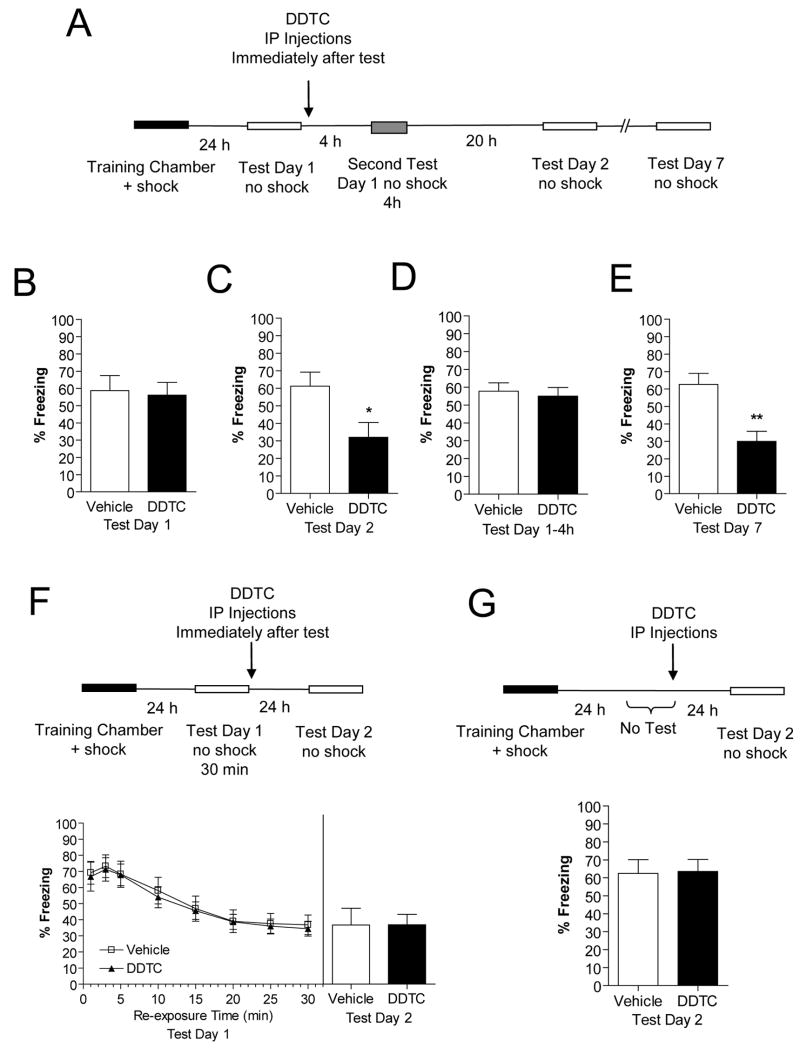
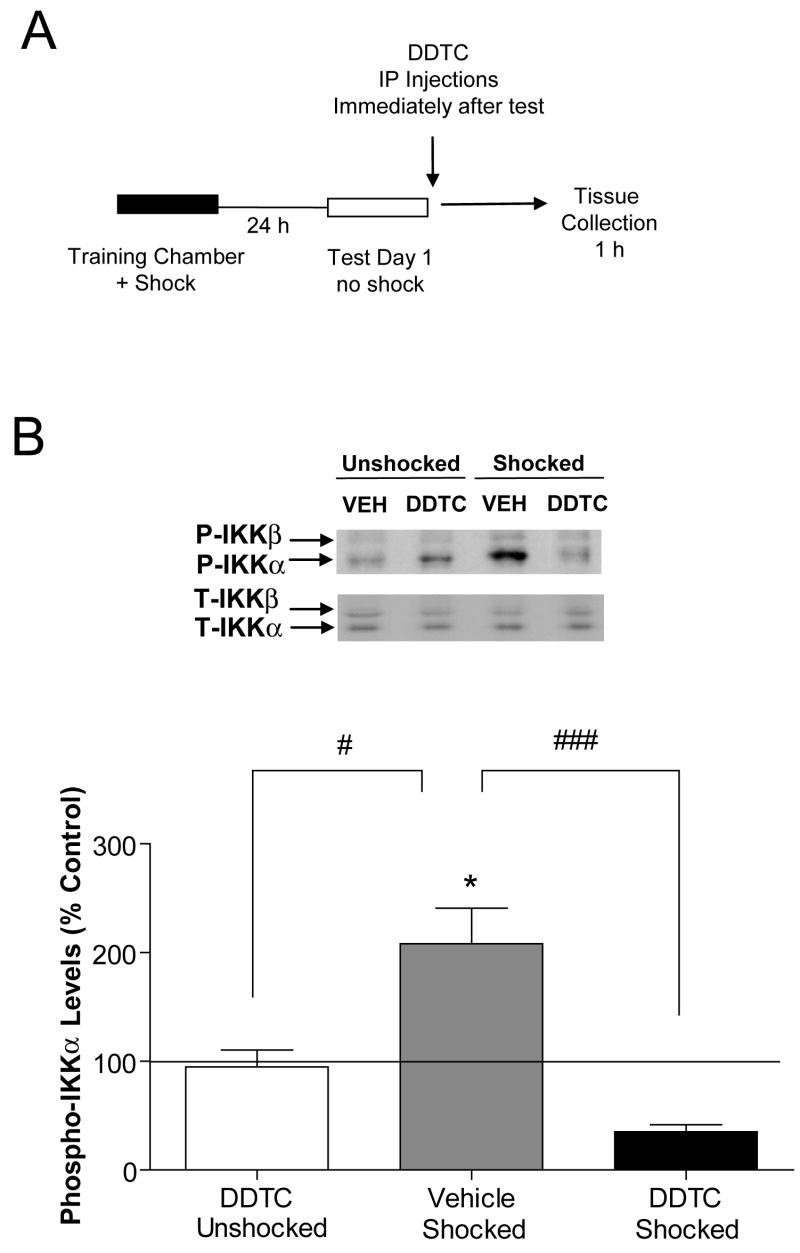
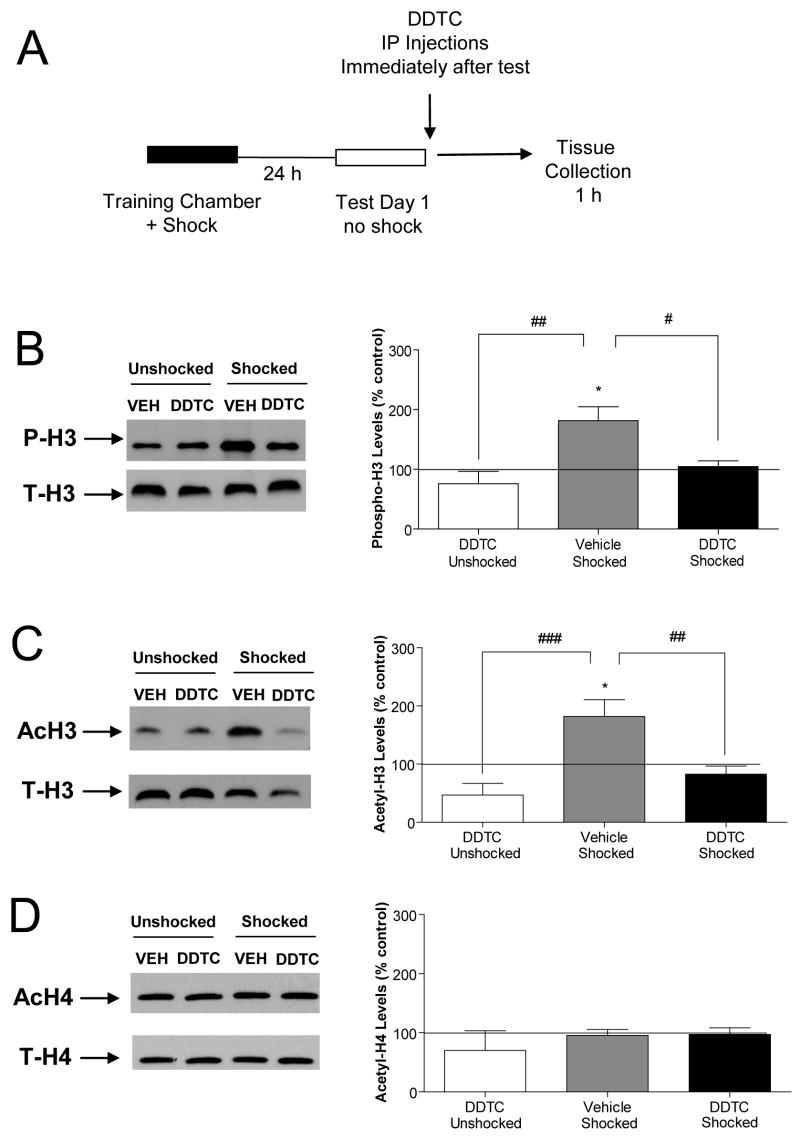
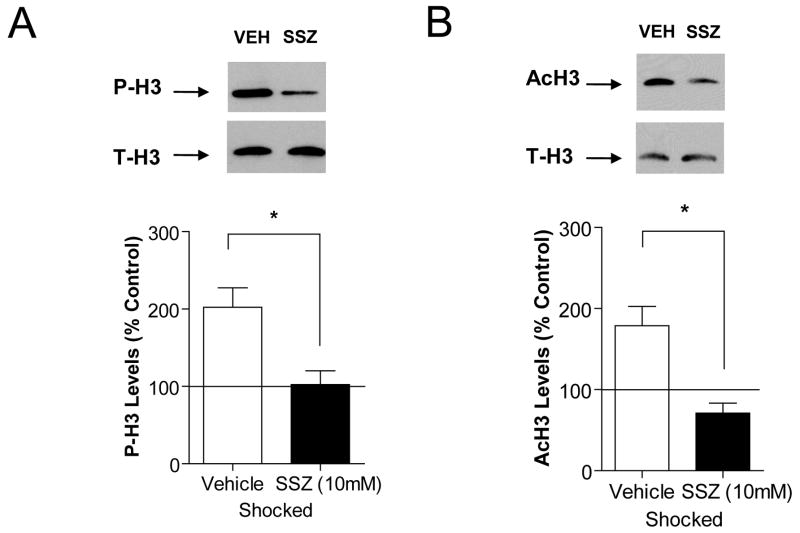
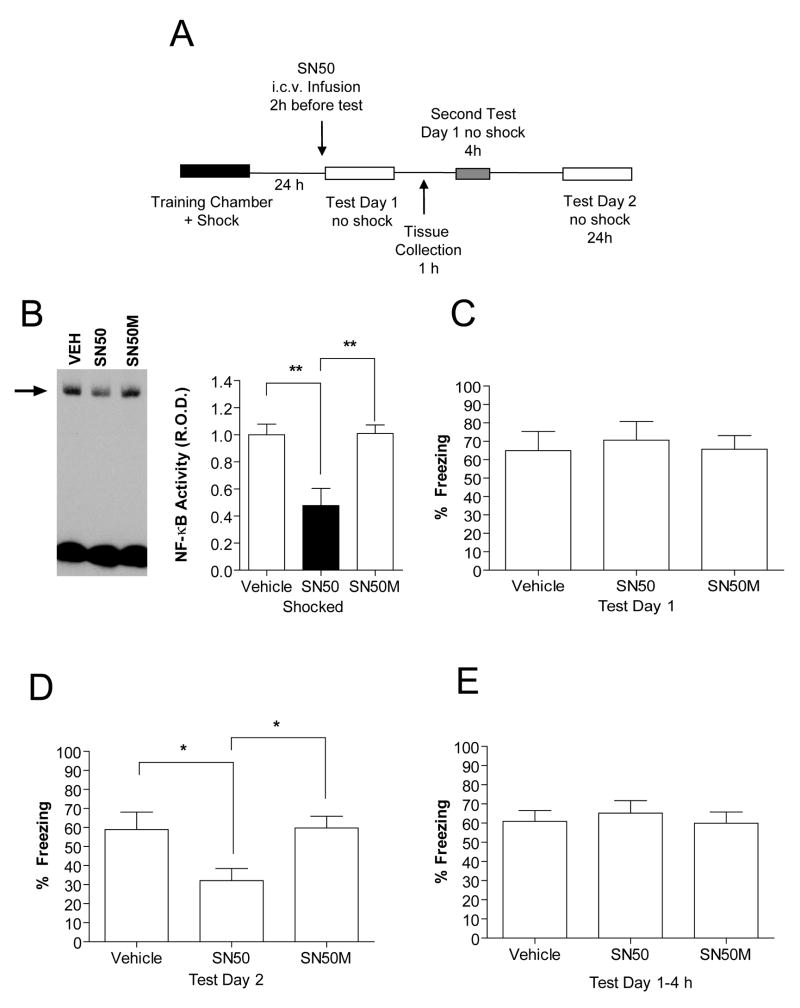
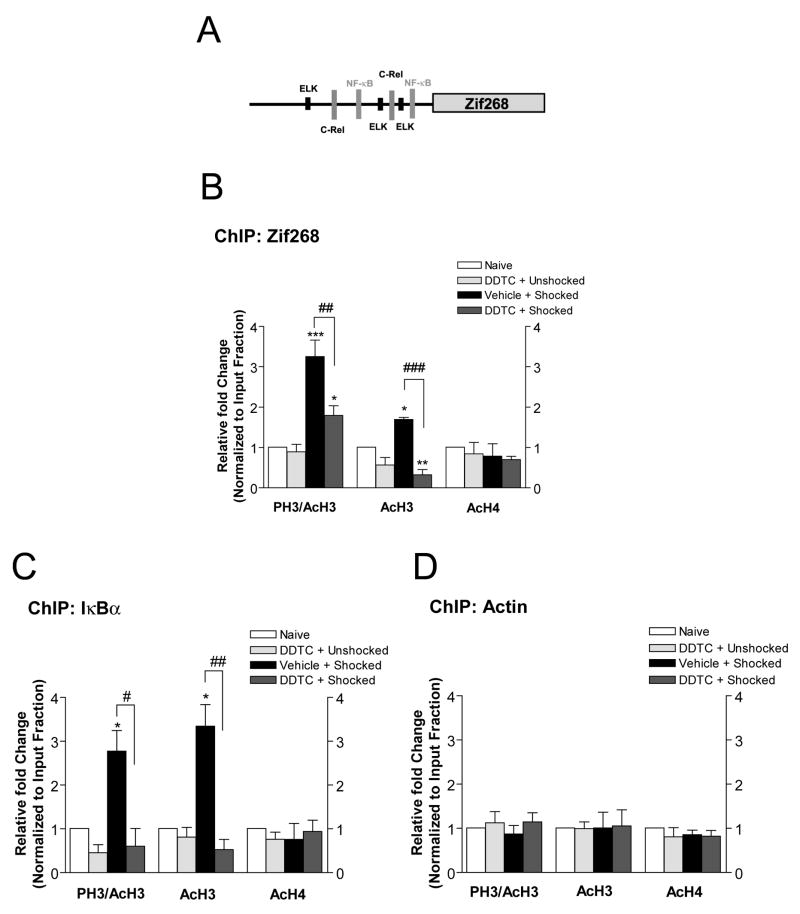
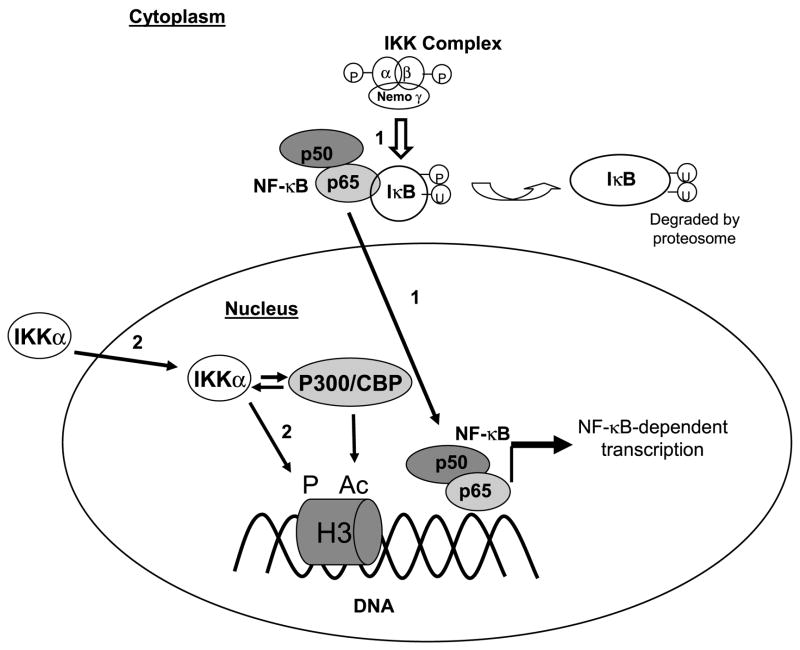
Previously formed memories are susceptible to disruption immediately after recall due to a necessity to be reconsolidated after retrieval. Protein translation mechanisms have been widely implicated as being necessary for memory reconsolidation, but gene transcription mechanisms have been much less extensively studied in this context. We found that retrieval of contextual conditioned fear memories activates the NF-kappaB pathway to regulate histone H3 phosphorylation and acetylation at specific gene promoters in hippocampus, specifically via IKKalpha and not the NF-kappaB DNA-binding complex. Behaviorally, we found that inhibition of IKKalpha regulation of either chromatin structure or NF-kappaB DNA-binding complex activity leads to impairments in fear memory reconsolidation, and that elevating histone acetylation rescues this memory deficit in the face of IKK blockade. These data provide insights into IKK-regulated transcriptional mechanisms in hippocampus that are necessary for memory reconsolidation.
Figures



References
-
- Alarcon JM, Malleret G, Touzani K, Vronskaya S, Ishii S, Kandel ER, Barco A. Chromatin acetylation, memory, and LTP are impaired in CBP+/− mice: a model for the cognitive deficit in Rubinstein-Taybi syndrome and its amelioration. Neuron. 2004;42:947–959. - PubMed
-
- Albensi BC, Mattson MP. Evidence for the involvement of TNF and NF-kappaB in hippocampal synaptic plasticity. Synapse. 2000;35:151–159. - PubMed
-
- Anest V, Cogswell PC, Baldwin AS., Jr IkappaB kinase alpha and p65/RelA contribute to optimal epidermal growth factor-induced c-fos gene expression independent of IkappaBalpha degradation. J Biol Chem. 2004;279:31183–31189. - PubMed
-
- Anest V, Hanson JL, Cogswell PC, Steinbrecher KA, Strahl BD, Baldwin AS. A nucleosomal function for IkappaB kinase-alpha in NF-kappaB-dependent gene expression. Nature. 2003;423:659–663. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
