

The impact of a boosting immunogen on the differentiation of secondary memory CD8+ T cells
- PMID: 17881444
- PMCID: PMC2169130
- DOI: 10.1128/JVI.01519-07
The impact of a boosting immunogen on the differentiation of secondary memory CD8+ T cells
Abstract
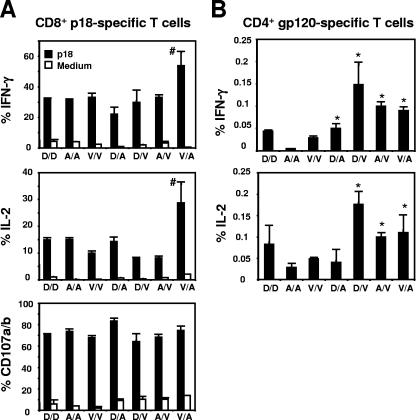
While recent studies have demonstrated that secondary CD8+ T cells develop into effector-memory cells, the impact of particular vaccine regimens on the elicitation of these cells remains poorly defined. In the present study we evaluated the effect of three different immunogens--recombinant vaccinia, recombinant adenovirus, and plasmid DNA--on the generation of memory cellular immune responses. We found that vectors that induce the rapid movement of CD8+ T cells into the memory compartment during a primary immune response also drive a rapid differentiation of these cells into effector-memory CD8+ T cells following a secondary immunization. In contrast, the functional profiles of both CD8+ and CD4+ T cells, assessed by measuring antigen-stimulated gamma interferon and interleukin-2 production, were not predominantly shaped by the boosting immunogen. We also demonstrated that the in vivo expression of antigen by recombinant vectors was brief following boosting immunization, suggesting that antigen persistence has a minimal impact on the differentiation of secondary CD8+ T cells. When used in heterologous or in homologous prime-boost combinations, these three vectors generated antigen-specific CD8+ T cells with different phenotypic profiles. Expression of the memory-associated molecule CD27 on effector CD8+ T cells decreased following heterologous but not homologous boosting, resulting in a phenotypic profile similar to that seen on primary CD8+ T cells. These data therefore suggest that the phenotype of secondary CD8+ T cells is determined predominantly by the boosting immunogen whereas the cytokine profile of these cells is shaped by both the priming and boosting immunogens.
Figures






Similar articles
-
Rapid memory CD8+ T-lymphocyte induction through priming with recombinant Mycobacterium smegmatis.J Virol. 2007 Jan;81(1):74-83. doi: 10.1128/JVI.01269-06. Epub 2006 Oct 18. J Virol. 2007. PMID: 17050608 Free PMC article.
-
A multigene HIV type 1 subtype C modified vaccinia Ankara (MVA) vaccine efficiently boosts immune responses to a DNA vaccine in mice.AIDS Res Hum Retroviruses. 2008 Feb;24(2):207-17. doi: 10.1089/aid.2007.0206. AIDS Res Hum Retroviruses. 2008. PMID: 18240963
-
Qualitative differences in cellular immunogenicity elicited by hepatitis C virus T-Cell vaccines employing prime-boost regimens.PLoS One. 2017 Jul 21;12(7):e0181578. doi: 10.1371/journal.pone.0181578. eCollection 2017. PLoS One. 2017. PMID: 28732046 Free PMC article.
-
Heterogeneity of CD4(+) and CD8(+) T cells.Curr Opin Immunol. 2003 Jun;15(3):336-42. doi: 10.1016/s0952-7915(03)00037-2. Curr Opin Immunol. 2003. PMID: 12787761 Review.
-
DNA vaccination: the answer to stable, protective T-cell memory?Curr Opin Immunol. 2003 Aug;15(4):471-6. doi: 10.1016/s0952-7915(03)00068-2. Curr Opin Immunol. 2003. PMID: 12900281 Review.
Cited by
-
Newborn mice vaccination with BCG.HIVA²²² + MVA.HIVA enhances HIV-1-specific immune responses: influence of age and immunization routes.Clin Dev Immunol. 2011;2011:516219. doi: 10.1155/2011/516219. Epub 2011 Apr 12. Clin Dev Immunol. 2011. PMID: 21603216 Free PMC article.
-
TLR1/2 activation during heterologous prime-boost vaccination (DNA-MVA) enhances CD8+ T Cell responses providing protection against Leishmania (Viannia).PLoS Negl Trop Dis. 2011 Jun;5(6):e1204. doi: 10.1371/journal.pntd.0001204. Epub 2011 Jun 14. PLoS Negl Trop Dis. 2011. PMID: 21695103 Free PMC article.
-
Sustained expression of alpha1-antitrypsin after transplantation of manipulated hematopoietic stem cells.Am J Respir Cell Mol Biol. 2008 Aug;39(2):133-41. doi: 10.1165/rcmb.2007-0133OC. Epub 2008 Mar 6. Am J Respir Cell Mol Biol. 2008. PMID: 18323534 Free PMC article.
-
Modulating numbers and phenotype of CD8+ T cells in secondary immune responses.Eur J Immunol. 2010 Jul;40(7):1916-26. doi: 10.1002/eji.201040310. Eur J Immunol. 2010. PMID: 20411564 Free PMC article.
-
Potentiating effects of MPL on DSPC bearing cationic liposomes promote recombinant GP63 vaccine efficacy: high immunogenicity and protection.PLoS Negl Trop Dis. 2011 Dec;5(12):e1429. doi: 10.1371/journal.pntd.0001429. Epub 2011 Dec 20. PLoS Negl Trop Dis. 2011. PMID: 22206029 Free PMC article.
References
-
- Bachmann, M. F., P. Wolint, K. Schwarz, P. Jager, and A. Oxenius. 2005. Functional properties and lineage relationship of CD8+ T cell subsets identified by expression of IL-7 receptor alpha and CD62L. J. Immunol. 175:4686-4696. - PubMed
-
- Haring, J. S., V. P. Badovinac, and J. T. Harty. 2006. Inflaming the CD8+ T cell response. Immunity 25:19-29. - PubMed
-
- Heath, W. R., and F. R. Carbone. 2001. Cross-presentation in viral immunity and self-tolerance. Nat. Rev. Immunol. 1:126-134. - PubMed
-
- Hendriks, J., L. A. Gravestein, K. Tesselaar, R. A. van Lier, T. N. Schumacher, and J. Borst. 2000. CD27 is required for generation and long-term maintenance of T cell immunity. Nat. Immunol. 1:433-440. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
