

FGF9 and SHH regulate mesenchymal Vegfa expression and development of the pulmonary capillary network
- PMID: 17881491
- PMCID: PMC2099314
- DOI: 10.1242/dev.004879
FGF9 and SHH regulate mesenchymal Vegfa expression and development of the pulmonary capillary network
Abstract
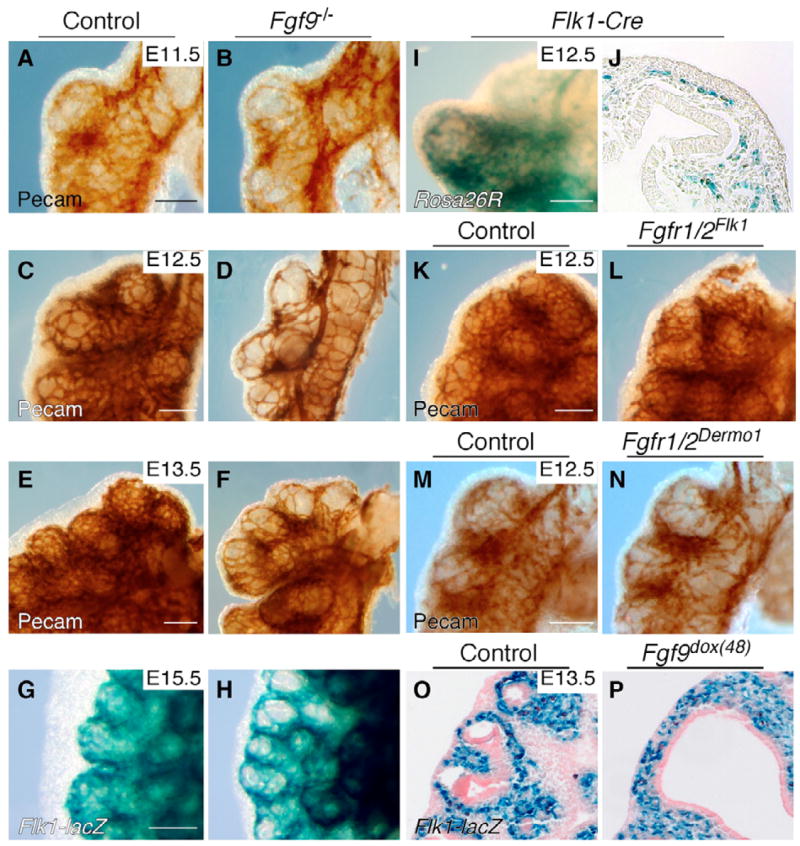
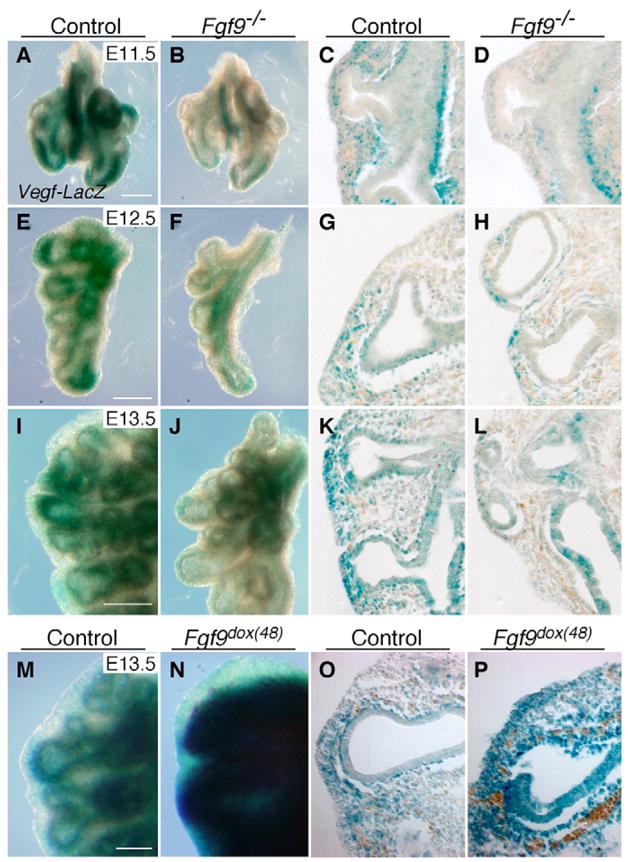
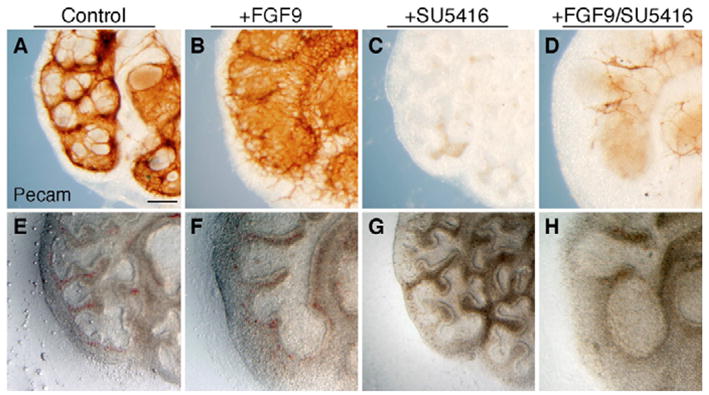
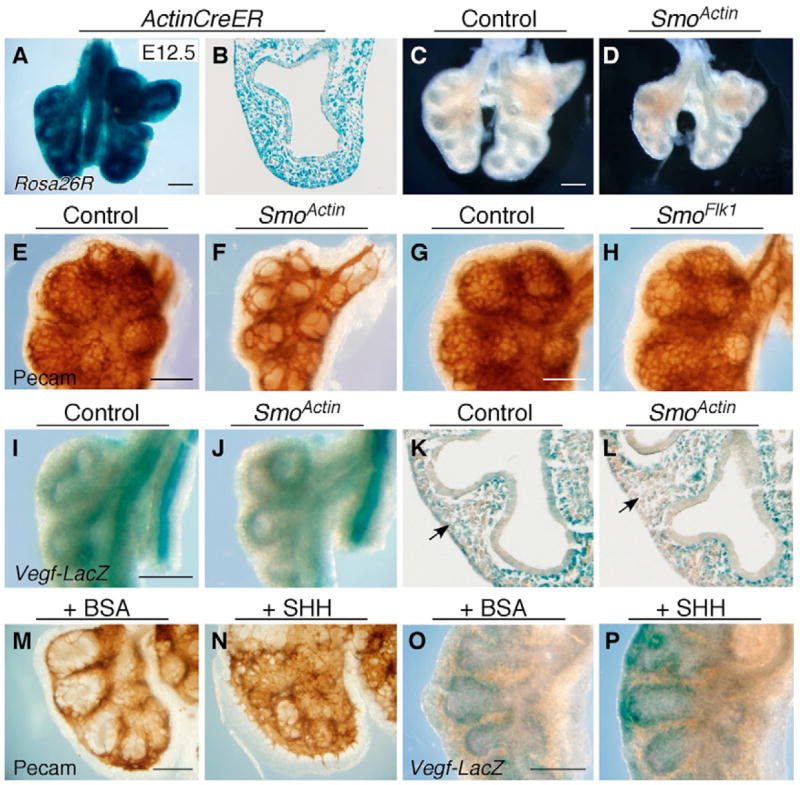
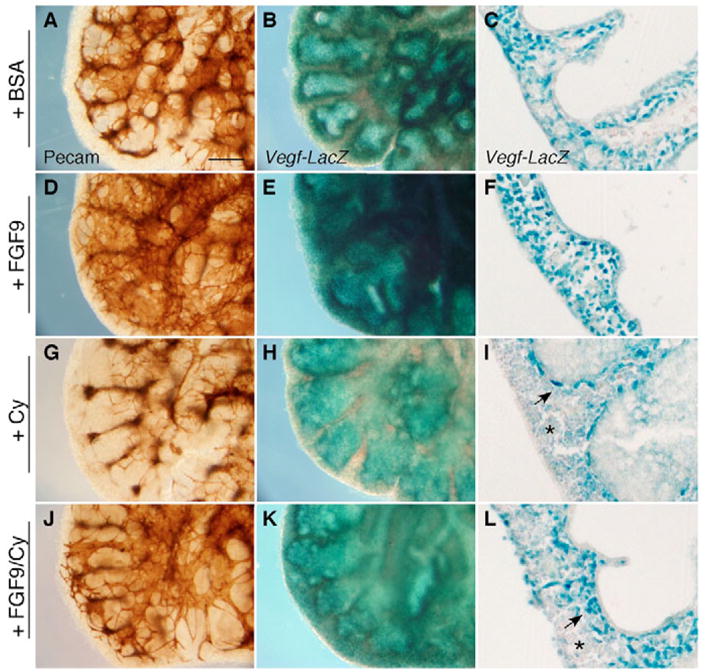
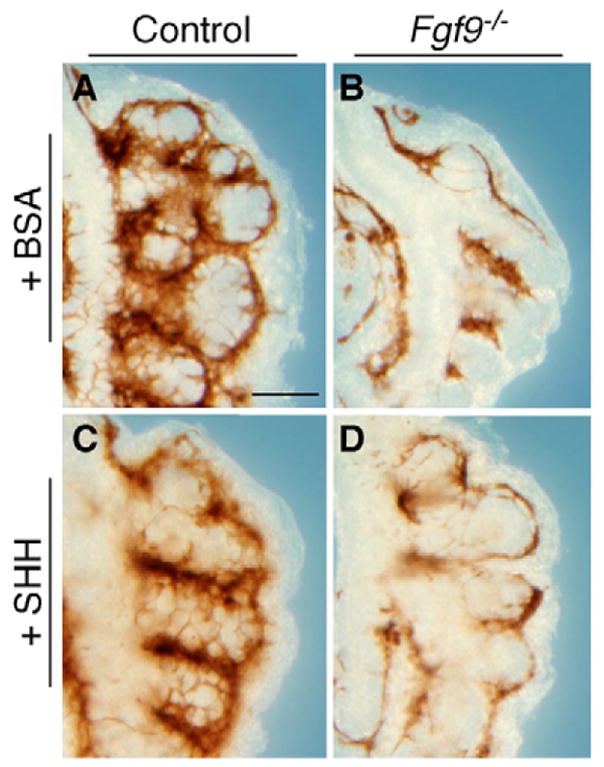
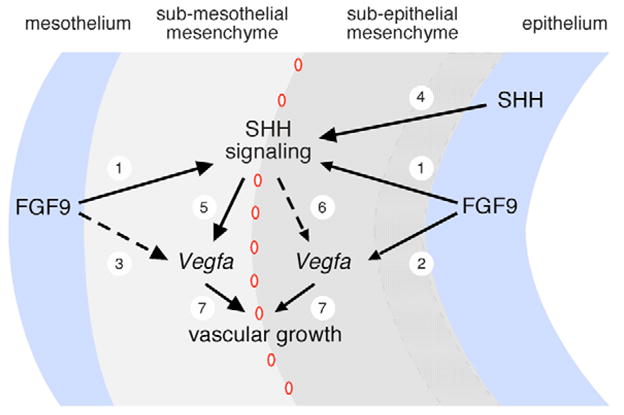
The juxtaposition of a dense capillary network to lung epithelial cells is essential for air-blood gas exchange. Defective lung vascular development can result in bronchopulmonary dysplasia and alveolar capillary dysplasia. Although vascular endothelial growth factor A (Vegfa) is required for formation of the lung capillary network, little is known regarding the factors that regulate the density and location of the distal capillary plexus and the expression pattern of Vegfa. Here, we show that fibroblast growth factor 9 (FGF9) and sonic hedgehog (SHH) signaling to lung mesenchyme, but not to endothelial cells, are each necessary and together sufficient for distal capillary development. Furthermore, both gain- and loss-of-function of FGF9 regulates Vegfa expression in lung mesenchyme, and VEGF signaling is required for FGF9-mediated blood vessel formation. FGF9, however, can only partially rescue the reduction in capillary density found in the absence of SHH signaling, and SHH is unable to rescue the vascular phenotype found in Fgf9(-/-) lungs. Thus, both signaling systems regulate distinct aspects of vascular development in distal lung mesenchyme. These data suggest a molecular mechanism through which FGF9 and SHH signaling coordinately control the growth and patterning of the lung capillary plexus, and regulate the temporal and spatial expression of Vegfa.
Figures
Similar articles
-
FGF9 and SHH signaling coordinate lung growth and development through regulation of distinct mesenchymal domains.Development. 2006 Apr;133(8):1507-17. doi: 10.1242/dev.02313. Epub 2006 Mar 15. Development. 2006. PMID: 16540513
-
Fibroblast growth factor 9 signaling inhibits airway smooth muscle differentiation in mouse lung.Dev Dyn. 2009 Jan;238(1):123-37. doi: 10.1002/dvdy.21831. Dev Dyn. 2009. PMID: 19097117 Free PMC article.
-
Reciprocal epithelial-mesenchymal FGF signaling is required for cecal development.Development. 2006 Jan;133(1):173-80. doi: 10.1242/dev.02175. Epub 2005 Nov 24. Development. 2006. PMID: 16308329 Free PMC article.
-
Growth factors in lung development.Adv Clin Chem. 2005;40:261-316. doi: 10.1016/s0065-2423(05)40007-4. Adv Clin Chem. 2005. PMID: 16355925 Review.
-
Sonic hedgehog signaling in the lung. From development to disease.Am J Respir Cell Mol Biol. 2015 Jan;52(1):1-13. doi: 10.1165/rcmb.2014-0132TR. Am J Respir Cell Mol Biol. 2015. PMID: 25068457 Free PMC article. Review.
Cited by
-
Cells responding to hedgehog signaling contribute to the theca of ovarian follicles.Reproduction. 2021 Apr;161(4):437-448. doi: 10.1530/REP-20-0471. Reproduction. 2021. PMID: 33720037 Free PMC article.
-
Pulmonary hypertension in the premature infant: a challenging comorbidity in a vulnerable population.Curr Opin Pediatr. 2016 Jun;28(3):324-30. doi: 10.1097/MOP.0000000000000355. Curr Opin Pediatr. 2016. PMID: 27043088 Free PMC article. Review.
-
Mesodermal lineages in the developing respiratory system.Trends Dev Biol. 2016;9:91-110. Trends Dev Biol. 2016. PMID: 34707332 Free PMC article.
-
FUBP3 interacts with FGF9 3' microsatellite and positively regulates FGF9 translation.Nucleic Acids Res. 2011 May;39(9):3582-93. doi: 10.1093/nar/gkq1295. Epub 2011 Jan 19. Nucleic Acids Res. 2011. PMID: 21252297 Free PMC article.
-
Implantation of fibrin gel on mouse lung to study lung-specific angiogenesis.J Vis Exp. 2014 Dec 21;(94):52012. doi: 10.3791/52012. J Vis Exp. 2014. PMID: 25548859 Free PMC article.
References
-
- Akeson AL, Greenberg JM, Cameron JE, Thompson FY, Brooks SK, Wiginton D, Whitsett JA. Temporal and spatial regulation of VEGF-A controls vascular patterning in the embryonic lung. Dev Biol. 2003;264:443–455. - PubMed
-
- Asahara T, Bauters C, Zheng LP, Takeshita S, Bunting S, Ferrara N, Symes JF, Isner JM. Synergistic effect of vascular endothelial growth factor and basic fibroblast growth factor on angiogenesis in vivo. Circulation. 1995;92:365–371. - PubMed
-
- Auguste P, Gursel DB, Lemiere S, Reimers D, Cuevas P, Carceller F, Di Santo JP, Bikfalvi A. Inhibition of fibroblast growth factor/fibroblast growth factor receptor activity in glioma cells impedes tumor growth by both angiogenesis-dependent and -independent mechanisms. Cancer Res. 2001;61:1717–1726. - PubMed
-
- Auguste P, Javerzat S, Bikfalvi A. Regulation of vascular development by fibroblast growth factors. Cell Tissue Res. 2003;314:157–166. - PubMed
-
- Bellusci S, Furuta Y, Rush MG, Henderson R, Winnier G, Hogan BL. Involvement of Sonic hedgehog (Shh) in mouse embryonic lung growth and morphogenesis. Development. 1997a;124:53–63. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
