

Egfr/Ras signaling regulates DE-cadherin/Shotgun localization to control vein morphogenesis in the Drosophila wing
- PMID: 17888420
- PMCID: PMC2128780
- DOI: 10.1016/j.ydbio.2007.08.003
Egfr/Ras signaling regulates DE-cadherin/Shotgun localization to control vein morphogenesis in the Drosophila wing
Abstract
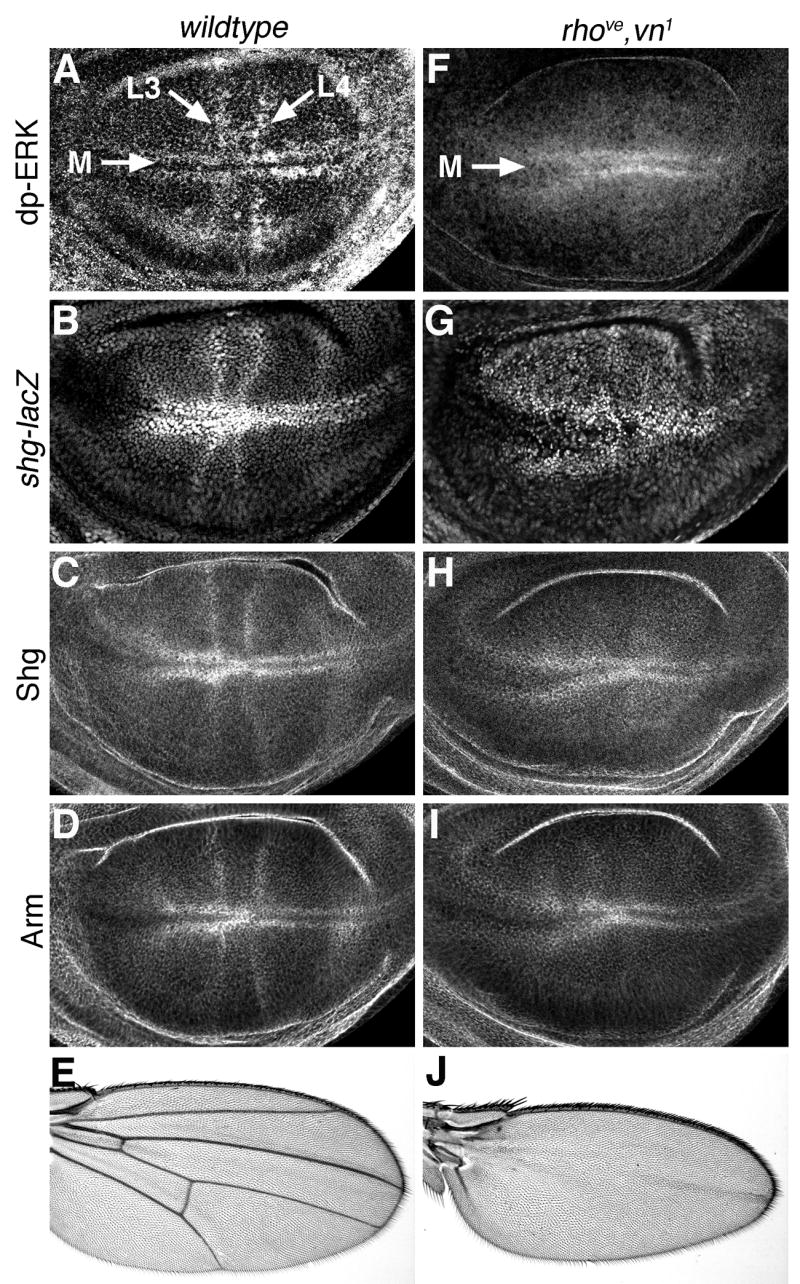
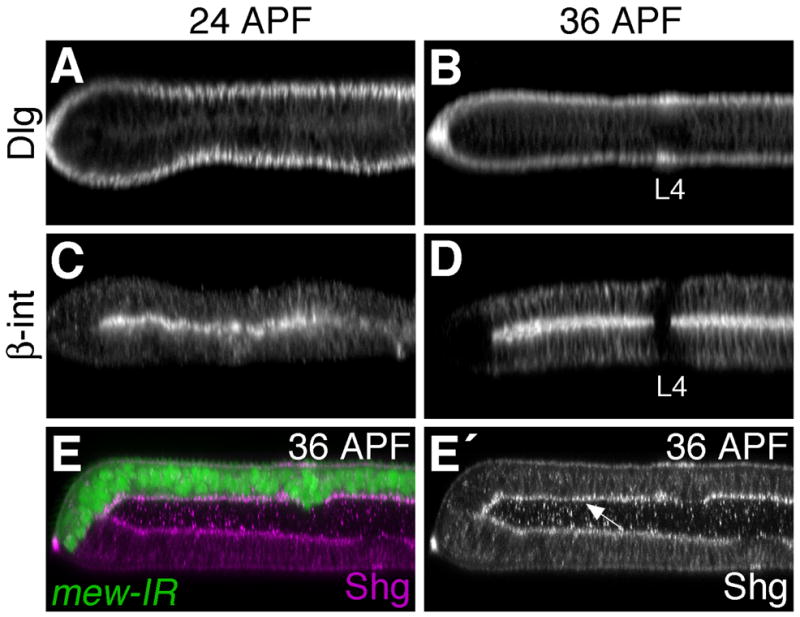
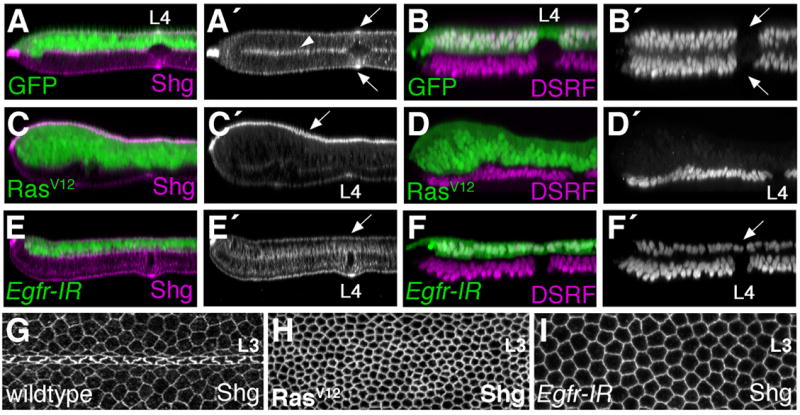
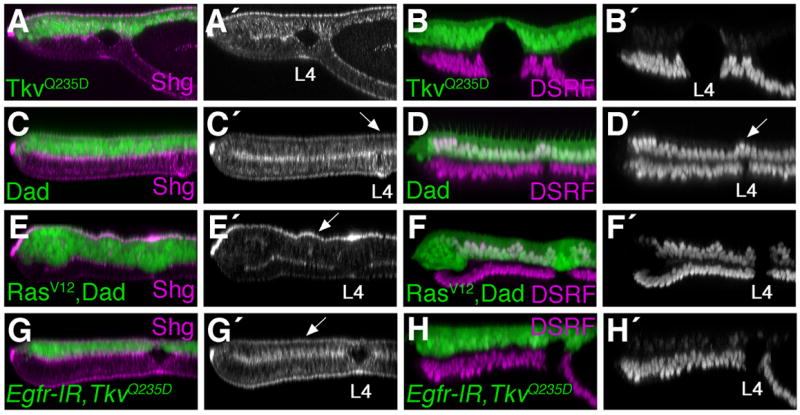
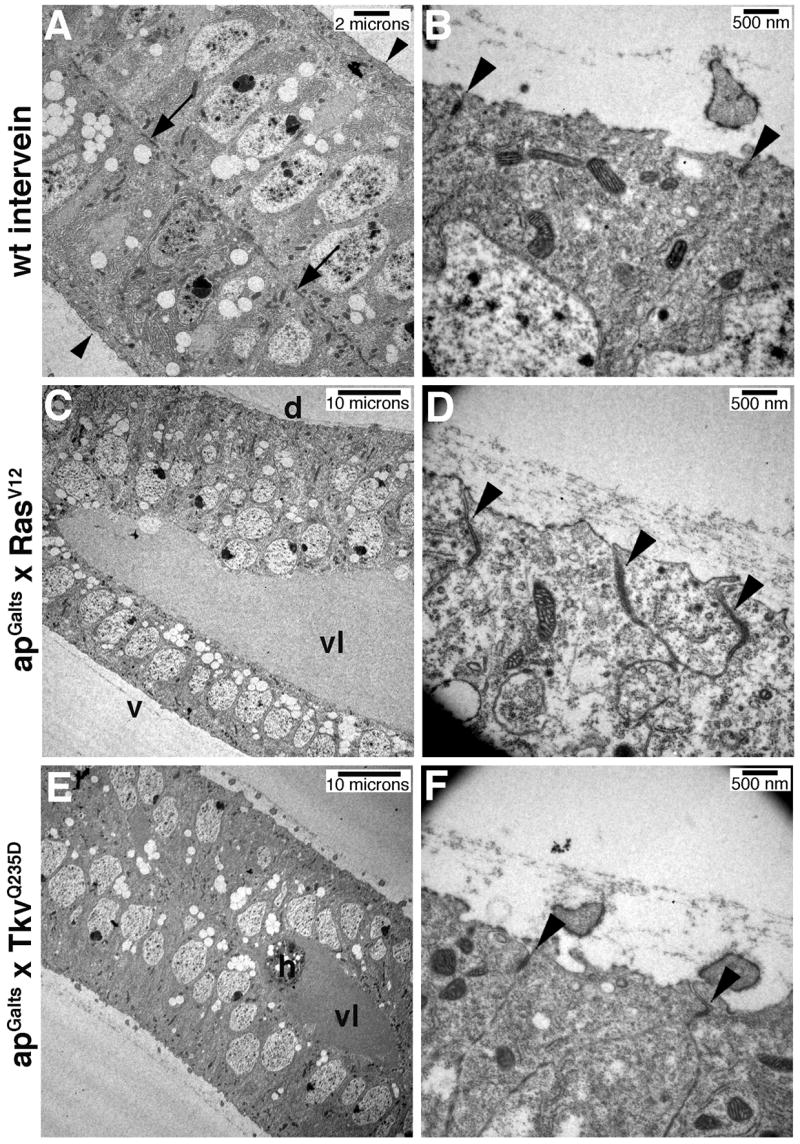
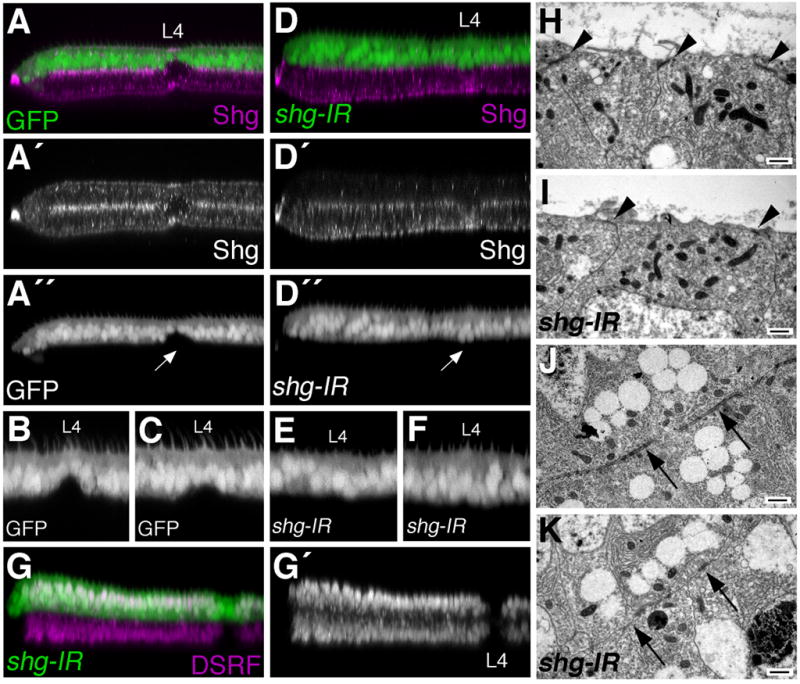
Egfr/Ras signaling promotes vein cell fate specification in the developing Drosophila wing. While the importance of Ras signaling in vein determination has been extensively documented, the mechanisms linking Ras activity to vein differentiation remain unclear. We found that Ras signaling regulates both the levels and subcellular localization of the cell adhesion molecule DE-cadherin/Shotgun (Shg) in the differentiating wing epithelium. High Ras activity in presumptive vein cells directs the apical localization of Shg containing adherens junctions, whereas low Ras activity in intervein cells allows Shg to relocalize basally. These alterations in Shg-mediated adhesion control cell shape changes that are essential for vein morphogenesis. While Decapentaplegic (Dpp) acts downstream of Ras to maintain vein cell identity in the pupal wing, our results indicate that Ras controls Shg localization via a Dpp-independent mechanism. Ras, therefore, regulates both the transcriptional responses necessary for vein cell identity, and the cell adhesive changes that determine vein and intervein cell morphology.
Figures



References
-
- Bier E. Localized activation of RTK/MAPK pathways during Drosophila development. Bioessays. 1998;20:189–94. - PubMed
-
- Brower DL, Jaffe SM. Requirement for integrins during Drosophila wing development. Nature. 1989;342:285–287. - PubMed
-
- Brown KE, Baonza A, Freeman M. Epithelial cell adhesion in the developing Drosophila retina is regulated by Atonal and the EGF receptor pathway. Dev Biol 2006 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
